Люстра клетка - Chandelier cell
| Люстра клетка | |
|---|---|
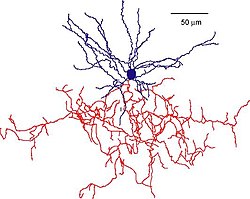 Реконструкция клетки люстры мышки. Сома и дендриты помечены синим цветом, ветви аксонов - красным. (Woodruff & Yuste, 2008, PLoS Biology).[1] | |
| Анатомические термины нейроанатомии |

Потенциал действия в пирамидном нейроне (клетка 1) вызывает спайк в клетке люстры (2) через сильную связь, в свою очередь, вызывая спайк третьего порядка в пирамидной клетке, расположенной ниже по течению (3). Этот всплеск приводит к регистрации трисинаптического ВПСП в постсинаптической пирамидной клетке (ячейка 4, событие А). В то же время ячейка 3 приводит к порогу как корзину-ячейку (5), так и ячейку люстры (6). Ячейка корзины вызывает гиперполяризационный IPSP на постсинаптически записанной пирамидной ячейке (ячейка 4, событие B), четыре синапса удалены из исходного шипа. Клетка люстры (6) запускает еще один пирамидальный нейрон (7), который производит ВПСП на зарегистрированном нейроне (ячейка 4, событие C), в пяти синапсах от исходного импульса. Результатом, наблюдаемым в постсинаптическом пирамидном нейроне (клетка 4), является задержанная последовательность EPSP-IPSP-EPSP (события A, B и C), проходящая через три, четыре и пять синапсов соответственно. Molnár et al. предлагать[2] что полисинаптические пути, подобные этому, могут быть активированы одним потенциалом действия в кортикальной пирамидной клетке.
Люстра нейроны или же люстры являются подмножеством ГАМКергический корковый интернейроны. Они описаны как парвальбумин -содержащий и быстрый-всплеск чтобы отличить их от других подтипов ГАМКергических нейронов, хотя более поздние исследования показали, что только подмножество люстрных клеток дает положительный результат на парвальбумин путем иммуноокрашивания.[3] Название происходит от специфической формы их аксон беседки, с выводами, образующими отдельные массивы, называемые "патроны". Картриджи иммунореактивны к изоформе мембранного транспортера ГАМК, ГАТ-1, и это служит их отличительной чертой.[4][5] GAT-1 участвует в процессе обратного захвата ГАМК нервными окончаниями, тем самым помогая прекратить его синаптическую активность. Синапс нейронов люстры исключительно к начальный сегмент аксона из пирамидные нейроны, рядом с участком где потенциал действия генерируется.[6] Считается, что они обеспечивают тормозящий эффект для пирамидных нейронов, но есть данные, показывающие, что в некоторых случаях ГАМК из нейронов люстр может быть возбуждающим. [7]
Картриджи аксонов, образованные клетками-канделябрами, являются одним из типов синапсов, которые демонстрируют самые драматические изменения в нормальном подростковом возрасте.[8] и потенциально может иметь отношение к началу психического заболевания у взрослых. Продолжая эту ссылку, в шизофрения, ученые наблюдали изменения в их форме и функциональности, например, снижение конечной плотности аксонов на 40%. [9]
внешняя ссылка
- SRF интервью Дэвида Льюиса - интервью, касающееся дисфункции ГАМКергических нейронов при шизофрении и роли люстр.
- Поиск NIF - Люстра Cell через Информационная структура по неврологии
- Корковое развитие - изображения нейронов люстры и информация об изменениях в их развитии. Программа трансляционной нейробиологии в Университете Питтсбурга.
- Как люстры освещают человеческую мысль - Тип клетки мозга, называемый нейроном люстры, может быть тем, что дает нам преимущество перед другими млекопитающими в мысли и языке, Новый ученый, 3 сентября 2008 г.[2]
Рекомендации
- ^ а б Вудрафф А., Юсте Р. (сентябрь 2008 г.). "О мышах, людях и люстрах". PLoS Biol. 6 (9): e243. Дои:10.1371 / journal.pbio.0060243. ЧВК 2553849. PMID 18816168.
- ^ а б Мольнар Г., Олах С., Комлози Г., Фюле М., Сабадич Дж., Варга С., Барзо П., Тамаш Г. (сентябрь 2008 г.). Юсте Р. (ред.). «Сложные события, инициированные отдельными спайками в коре головного мозга человека». PLoS Biol. 6 (9): e222. Дои:10.1371 / journal.pbio.0060222. ЧВК 2528052. PMID 18767905.
- ^ Танигучи Х., Лу Дж., Хуанг З. Дж. (Январь 2013 г.). «Пространственное и временное происхождение люстр в неокортексе мыши». Наука. 339 (6115): 70–4. Bibcode:2013Наука ... 339 ... 70 т. Дои:10.1126 / science.1227622. ЧВК 4017638. PMID 23180771.
- ^ Борден, Л. А. (1996). «Гетерогенность транспортера ГАМК: фармакология и клеточная локализация». Международная нейрохимия. 29 (4): 335–356. Дои:10.1016/0197-0186(95)00158-1. PMID 8939442. S2CID 25089539.
- ^ Hardwick, C .; French, S.J .; Southam, E .; Тоттерделл, С. (2005). «Сравнение возможных маркеров для патронов люстр в медиальной префронтальной коре и гиппокампе крыс». Исследование мозга. 1031 (2): 238–244. Дои:10.1016 / j.brainres.2004.10.047. PMID 15649449. S2CID 25594278.
- ^ Контрерас, Д. (2004). «Электрофизиологические классы нейронов неокортекса». Нейронные сети. 17 (5–6): 633–646. Дои:10.1016 / j.neunet.2004.04.003. PMID 15288889.
- ^ Szabadics, J .; Varga, C .; Molnár, G .; Oláh, S .; Barzó, P .; Тамаш, Г. (2006). «Возбуждающее действие ГАМКергических аксо-аксонных клеток в корковых микросхемах». Наука. 311 (5758): 233–235. Bibcode:2006Научный ... 311..233S. Дои:10.1126 / science.1121325. PMID 16410524. S2CID 40744562.
- ^ Anderson, S.A .; Classey, J.D .; Конде, Ф .; Lund, J.S .; Льюис, Д.А. (1995). «Синхронное развитие дендритных шипов пирамидных нейронов и парвальбумин-иммунореактивных терминалов аксонов нейронов люстр в слое III префронтальной коры головного мозга обезьян». Неврология. 67 (1): 7–22. Дои:10.1016 / 0306-4522 (95) 00051-J. ISSN 0306-4522. PMID 7477911. S2CID 25474218.
- ^ Pierri, J. N .; Chaudry, A. S .; Woo, T. U .; Льюис, Д. А. (1999). «Изменения в терминалах аксонов нейронов люстры в префронтальной коре головного мозга шизофреников». Американский журнал психиатрии. 156 (11): 1709–1719. Дои:10.1176 / ajp.156.11.1709 (неактивно 18.10.2020). PMID 10553733.CS1 maint: DOI неактивен по состоянию на октябрь 2020 г. (связь)