Южнополярный регион мелового периода - South Polar region of the Cretaceous

Было предложено, чтобы эта статья была слился в Меловой. (Обсуждать) Предлагается с ноября 2020 года. |
В Южнополярный регион мелового периода состоит из континента Восточная Гондвана–Современная Австралия и Антарктида – продукт распада Гондвана. Южный регион в то время был намного теплее, чем сегодня, и колебался от 4–8 ° C (39–46 ° F) в самый поздний меловой период. Маастрихтский на территории нынешней юго-восточной Австралии. Это препятствовало развитию постоянных ледовых щитов и способствовало развитию полярные леса, в которых преобладали хвойные породы, саговники, и папоротники, и полагался на умеренный климат и проливные дожди. Основные ископаемые геологические образования которые записывают эту область: Санта-Марта и Собральские образования из Остров Сеймур от Антарктический полуостров; в Сноу-Хилл-Айленд, Лопес де Бертодано, а Скрытые образования озера на Остров Джеймса Росса также у Антарктического полуострова; и Eumeralla и Формации Wonthaggi в Австралии.
В Южном полярном регионе проживало много эндемичный виды, в том числе несколько реликт формы, которые вымерли в других местах к меловому периоду. Из множества динозавров наиболее разнообразными были мелкие гипсилофодонт -подобные динозавры. Южнополярный регион также был домом для последних лабиринтодонт амфибия Koolasuchus. Изоляция Антарктиды породила особую экосистему морской жизни, названную провинцией Уэдделлиан.
Пейзаж
Геология

Бассейн Латади на юге Антарктического полуострова -Палмер Лэнд -содержит вулканическая порода, свидетельствуя о большом магматический событие в Средний мел называемое событием Палмера Лэнда. Эта область дала множество макро- и микро- завод и моллюск ископаемые представители Раннемеловой период, так же хорошо как Середина к Поздняя юра.[2][3] Район, который сейчас является Антарктическим полуостровом, на протяжении мезозоя был подчинение прототихоокеанский пластина, вызывая вулканическую активность.[4]
Важный ископаемый формирование это Формация Санта-Марта в Остров Джеймса Росса от Антарктический полуостров, одно из самых значительных образований Южное полушарие, представляющий большое разнообразие наземной флоры и фауны Поздний мел Полярный регион. Остров Сеймур аналогична в этом отношении область открытий для позвоночное животное и беспозвоночный существа - такие как плезиозавры –– позднего мела, простираясь в эоцен.[3] Другие меловые ископаемые образования в Антарктиде - это Формация острова Сноу-Хилл, то Формация Лопес де Бертодано, а Формация Скрытого озера, также на острове Джеймса Росса.[5][6]
В Формация Эумералла из Бухта динозавров и Формация Вонтхагги в Виктории, Австралия, содержат ряд окаменелостей динозавров и треки из раннего мелового периода. В Формация Тахора представляет собой совокупность рептилий меловой Новой Зеландии.[6]
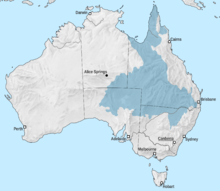
Море Эроманга было внутреннее море через территорию Австралии, сформировавшейся в раннем меловом периоде. Море достигло Бассейн Эроманга с севера через Карпентарианский бассейн. В южной части моря находились лагуны и реки, а на востоке - в Суратская котловина бухта. Когда Индия отошла от Австралии в раннем меловом периоде, Пертский бассейн также наполнен морской водой. Область, которая будет Коралловое море был рифтовая долина. Когда Австралия и Антарктида отделились на протяжении мелового периода, между ними образовалось море.[7]
Растительность
В Меловой характеризуется высокими глобальными температурами, вызванными большим количеством углекислый газ и возможно метан парниковые газы в атмосфере. Это вызвало отсутствие постоянного ледового покрова в полярных регионах, хотя уровень углекислого газа упал между 115 и 66 миллион лет назад (млн лет назад), что, возможно, дает постоянный ледяной покров. Возможно, что на протяжении мелового периода образовалось несколько небольших ледниковых щитов.[8] Повысился тектоническая активность, вызывая больше геотермальное тепло, возможно, предотвратили оледенение и повышение глобальной температуры. Температура могла быть на 15 ° C (27 ° F) выше, чем в наше время.[9][10][11]
Юрский
Пейзаж Средняя юра Полярный регион был реконструирован по остаткам лесов, обнаженных в Новой Зеландии, которые находились между 70 и 80 ° ю.ш. в юрский период. Растительность в основном состояла из хвойных деревьев, саговники, и другие голосеменные, а также папоротники; на лесной подстилке рос ликоподы, мохообразные, грибы и водоросли. Он имел умеренный климат с обильными дождями.[12][13]
Раннемеловой период
в Раннемеловой период, Восток Гондвана (Австралия, Антарктида и Зеландия ) начал отделяться от Южной Америки, и Индия и Мадагаскар также начали отделяться примерно в то же время.[8] В тропический зона могла расшириться в меловом периоде до 32 ° ю.ш., что позволяет деревьям в Антарктике расти круглый год в полярные леса. Открытие нескольких зрелых вечнозеленый и лиственный деревья указывают на температуру от теплой до прохладной с умеренным сезоном без повсеместных замерзаний, по крайней мере, между широты 70 и 85 ° ю.ш..[14][15] Однако также возможно, что жизнь растений может быть репрезентативной только для теплых летних месяцев.[16] Многое из того, что известно о растительной жизни Восточной Гондваны в меловой период, состоит из пыльца остается и сжатие листьев с севера Антарктический полуостров.[17] В зависимости от широты полярные зимы могло длиться от шести недель до четырех с половиной месяцев.[18]

Однако породы раннего мела Формация Вонтхагги на юго-востоке Австралии есть свидетельства сезонной мерзлоты.[20] Его географическое положение в раннем меловом периоде на отметке 78 ° ю.ш. указывает на то, что в этом районе зимой было темно от одного до трех месяцев, и этот район является типичным для ледникового питания. пойма.[21] Свидетельства раннего мелового периода оледенение и холодный климат был обнаружен в отложениях в Бассейн Эроманга в современной центральной Австралии, или от 60 до 80 ° ю. ш. в раннем меловом периоде. Этот бассейн был большим внутреннее море в меловом периоде. Тем не менее, подобные образования могли быть созданы просто потоком селей, и поэтому вполне возможно, что оледенения там никогда не было.[22][23] Общая площадь полярного льда во время Мезозойский возможно, был на треть меньше, чем в наше время, хотя похолодание минусовых температур, возможно, имели место на протяжении всего раннего мела.[24]
Поздний мел
В навес полярных лесов вокруг современного Остров Александра, который был вокруг 75 ° ю.ш. в среднем меловом периоде были преимущественно вечнозелеными, и, вероятно, большинство южнополярных лесов также были и состояли в основном из араукарианский и подокарпий хвойные породы.[3][25][15] Считается, что эти деревья остались бездействующий всю полярную зиму до лета под полуденное солнце.[26]
Доказательство того цветущие растения возрастом около 80 млн лет в Поздний мел предполагает наличие умеренные леса - такие же, как в современной Австралии, Новой Зеландии и южной части Южной Америки.[19] Некоторые остатки цветов были обнаружены рядом 60 ° ю.ш., и возможно на такой низкой широте, что эта область была подвержена полярным зимам и сезонной погоде, хотя цветы предполагают годовой диапазон температур 8–15 ° C (46–59 ° F) и дождливый климат.[15][17]
Остатки пыльцы из юго-востока Австралии идентичны живым видам растений Австралии: хвойным, цветущим растениям, населяющим районы с большим количеством осадков и средиземноморский климат, и склерофильный кустарники, что указывает на уникальный ландшафт тропических лесов и открытых кустарников.[27] Обозначается размером годичные кольца на ископаемых деревьях в полярных лесах Антарктики на протяжении всего Маастрихтский возраст 72–66 миллионов лет назад, от средней годовой температуры 7 ° C (45 ° F) до более сезонной экстремальной температуры 4–8 ° C (39–46 ° F).[28] Эти растения, вероятно, пережили Меловое – палеогеновое вымирание, который убил большую часть жизни 66 млн лет на вулканической Антарктический полуостров; окаменелости растений, датируемые 60 млн лет назад в Палеоцен из Остров Сеймур у берегов этого полуострова считаются предками растений умеренного пояса, населяющих современную Австралию и Южную Америку.[26]
Поздний мел (Кампанский ) Замек и Половина трехочковых формаций из Остров Кинг Джордж были расположены на 60 ° южной широты и отображают богатый комплекс ископаемой флоры, такой как Подокарпус; Араукария; в лептоспорангиатные папоротники Cladophlebis и Clavifera; и множество Магнолиопсида цветущие растения, Дикотилофиллум, Myrciophyllum santacruzensis, Нотофагус, Sterculiaephyllum australis, Монимиофиллум, и так далее.[29][30][31] В Формация Собрал острова Сеймур на границе мела и палеогена на палеошироте 63 ° ю. предоставил новый род ископаемых цветов в семье Cunoniaceae, Eucryphiaceoxylon eucryphioides.[32]
Экология
Как и в сегодняшней Австралии, Восточная Гондвана принимала многих эндемичный животных, среди которых много реликт виды семейств, которые вымерли в остальной части мелового мира. Возможно, полярные регионы позднего мела были заселены группами растений и животных, происхождение которых можно проследить до Ордовик.[15][33] Постепенная изоляция Антарктиды в позднем меловом периоде создала особую группу водных существ, получившую название Провинция Уэдделлиан.[34]
Динозавры
Птицы

Останки предка современных птиц, Neornithes, необычны в мезозое, с большим радиация происходящее в Неоген Антарктиды. Однако открытие позднего мелового периода Вегавис, а Гусь -подобная птица, на Остров Вега указывает на то, что основные современные группы птиц были обычны уже в меловом периоде. Бедренная кость неопознанного серия -подобная птица была обнаружена и на острове Вега. Следы птиц сохранились в бухте динозавров и, будучи крупнее большинства видов меловых птиц, указывают на обилие более крупных птиц. энантиорнит или же орнитурин птицы в раннем меловом периоде.[35]
Две ныряющие птицы, возможно примитивные гагары, были обнаружены в позднемеловом Чили и Антарктиде: Neogaeornis и Polarornis. Polarornis возможно, был способен как нырять, так и летать.[35] Раннее пингвины, Crossvallia и Вайману, известны от 61–62 млн лет назад в г. Палеоцен Однако молекулярные данные предполагают, что пингвины впервые появились в позднем меловом периоде. Учитывая, что эти пингвины были датированы так близко к событию мелового – палеогенового вымирания, группа эволюционировала либо до этого события, либо очень быстро после него.[35][36]
Нептичьи

Окаменелости динозавров в Южном полярном регионе редки, и основными местами хранения окаменелостей являются Группа острова Джеймса Росса; Ледник Бердмор в Антарктиде; Рома, Квинсленд; Ручей Мангахуанга в Новой Зеландии; и Бухта динозавров в Виктория, Австралия.[38] Динозавр остатки этого региона, такие, как те, что в Виктории, состоит только фрагментарные части, что делает идентификацию спорной. Например, оспариваемые идентификационные данные аллозаврид который может представлять Абелизаврид, то цератопсий Серендипацератопс который мог бы быть анкилозавр, и трудно классифицируемый теропод Тимимус следовательно, были сделаны.[37]
Суперконтинент Пангея юрского периода позволил крупным кладам динозавров достичь глобального распространения до распада, и между южнополярными формами и формами, встречающимися в других местах, существовало несколько близкородственных родственников, несмотря на разделение Тетис Океан. Однако группы динозавров, которые достигли пангондванского распространения в течение мелового периода, должны были использовать наземный мост, соединяющий Австралию с Южной Америкой через Антарктиду в Южном полярном регионе.[33] Южный полярный игуанодонт Муттабурразавр наиболее близок к европейскому рабдодонтиды, которые были доминирующей группой в Европе в позднем меловом периоде. Южный полярный меловой период Кунбарразавр определяется как самый базальный (примитивный) анкилозавр, что важно, поскольку анкилозавры известны как в Гондване, так и в Лавразии.[33][39] В Dromaeosauridae известны из Антарктиды и представляют собой реликтовую популяцию, ранее распространенную по всему миру.[40] Несмотря на эти очевидные межконтинентальные миграции, маловероятно, что южнополярные динозавры мигрировали из полярных лесов зимой, поскольку они были либо слишком массивными, как анкилозавры, либо слишком маленькими, например троодонтиды - путешествовать на большие расстояния, а большое море между Восточной Гондваной и другими континентами препятствовало любым таким миграциям в позднем меловом периоде.[41][42] Вполне возможно, что некоторые динозавры справятся с зимними условиями. впал в спячку, такой как теропод Тимимус.[43]

На сегодняшний день наиболее распространенной и разнообразной группой являются гипсилофодонт -подобные динозаврам, составляющие половину таксонов динозавров, обнаруженных в юго-восточной Австралии, что невидимо в более тропических регионах, что, возможно, указывает на какое-то преимущество перед другими динозаврами на полюсах.[15] Будучи маленькими с точеными зубами, они, вероятно, питались низменной растительностью, такой как ликоподы и подокарпии. стручки семян.[44] Гипсилофодонтовидный Leaellynasaura имел большой глазницы, крупнее, чем тропические динозавры, похожие на гипсилофодонтов, и, возможно, ночное видение, предлагая, что Leaellynasauraи, возможно, другие подобные гипсилофодонтам динозавры жили в полярных районах круглый год или большую часть года, включая полярные зимы. Костный рост был непрерывным на протяжении всей его жизни, что указывало на то, что он не впадал в спячку, возможно, если он эндотермический или же пойкилотермный,[38][45] или рыть норы.[46] Однако не исключено, что большие глаза возникли просто из-за онтогенез, то есть относительно большие глазницы могли быть признаком только молодых особей или, возможно, были врожденным дефектом, поскольку известен только один экземпляр.[41][18]
Хотя останки скудны и, следовательно, таксономические описания могут быть сомнительными,[47] останки викторианских теропод были отнесены к семи разным кладам: Цератозаврия, Спинозаврия, Тираннозавройда, Манираптора, Орнитомимозаврия, и Аллозавройда. Однако тираннозавроиды не известны на других континентах Гондваны, а более известны на севере Лавразия.[39] В отличие от других континентов Гондваны, чьи высшие хищники где абелизавриды и кархародонтозавриды, открытие Австраловенатор, Rapator и безымянный вид в Австралии предполагает мегарапторанс были главными хищниками Восточной Гондваны. Хвостовые позвонки неизвестного теропода, получившего название "Теропод Джоан Виффен ", были обнаружены в Поздняя юра / Раннемеловые породы Новой Зеландии.[48]

Три титанозавры –Саванназавр, Диамантиназавр, и Винтонотитан -и один макронарий –Австрозавр –Обнаружен в Формация Винтон составить зауропод Однако в сообществе меловой Австралии эти существа, вероятно, избегали полярных регионов, поскольку их останки полностью отсутствуют в юго-восточной Австралии, которая находилась в южнополярном регионе в меловом периоде. Однако вполне вероятно, что по крайней мере титанозавры мигрировали в Австралию из Южной Америки, что потребовало бы их прохождения через Антарктиду, поскольку титанозавры эволюционировали в меловом периоде после распада Пангеи. Возможно, событие Бонарелли в среднем меловом периоде сделало Антарктиду теплее и, следовательно, более гостеприимной для зауроподов.[39] Эти динозавры, вероятно, питались мясистыми семенами подокарпа и тис деревья, а также банальное раздвоенные папоротники времени.[44] Австрозавр может представлять реликт зауроподов средней юры, которые кажутся более примитивными, чем зауроподы мелового периода; неизвестно, почему более примитивные зауроподы пережили более производных зауроподов. Возможно, что динозавры, вымершие на экваторе, более предпочитали полярные регионы.[33]
Палеоцен
После астероид удар, последующий ударная зима считается, что убил динозавров вместе с большей частью мезозойской жизни в Меловое – палеогеновое вымирание. Однако отсутствие резкого вымирания горизонт в антарктических или австралийских отложениях для растений и двустворчатый окаменелости в этот период времени указывают на менее мощное воздействие в Южнополярном регионе.[15] Учитывая, что динозавры и другая фауна полярных регионов мелового периода были хорошо приспособлены к жизни в длительные периоды темной и холодной погоды, было высказано предположение, что это сообщество могло пережить это событие.[18]
Реки и озера
Последний темноспондилы –Группа гигантов амфибии которые в основном вымерли после Триасовый - населял Южный полярный регион в раннем меловом периоде. Koolasuchus, возможно, последний из темноспондилов, как полагают, выжили в регионах, где было слишком холодно для их конкурентов, неозухийцы –Группа рептилий, содержащая современные крокодилы - которые неактивны в воде при температуре ниже 10 ° C (50 ° F), чтобы выжить.[49] Хотя неозухийцы известны из Австралии мелового периода, считается, что они держались подальше от полярного региона и прибывали в Австралию по морю, а не по суше.[33]

Вероятно, темноспондили населяли пресноводные системы полярной Австралии до Бонарелли Событие в среднем меловом периоде около 100 млн лет назад температура повысилась, что позволило неозухиям заселить Антарктиду. Эти неозухианы при росте взрослой особи не более 2,5 метра (8,2 фута) в длину, вероятно, привели к исчезновению темноспондилов в тандеме с более развитыми. рыба с плавниками которые, возможно, были нацелены на их личинок. Миграция неозухийцев в регион предполагает, что средние зимние температуры были выше 5,5 ° C (41,9 ° F), а среднегодовая температура - более 14,2 ° C (57,6 ° F). Однако полярные неозухианы известны только по почти полному скелету Исисфордия, и другие остатки неозухий неопределенного вида.[49]
Плезиозавры населяли пресноводные реки и эстуарии, учитывая местонахождение их останков, вероятно, колонизировали Австралию в ранне-средней юре. Их останки, в основном зубы, были зарегистрированы на юго-востоке Австралии и относятся к позднему меловому периоду, хотя они никогда не были описанный так как останки слишком малочисленны для этого. Зубы имеют некоторое сходство с плиозавры, в частности ромалеозавриды и Leptocleidus, который вымер в раннем меловом периоде, что указывает на то, что полярные пресноводные системы могли быть убежищем для плиозавров мелового периода. В отличие от современных морских рептилий, эти южнополярные плезиозавры, вероятно, лучше переносили более холодные воды.[50][23]
Океаны

Ранний и средний мел морская рептилия остатки Южной Австралии включают пять семьи плезиозавров–Cryptoclididae, Elasmosauridae, Поликотилиды, Rhomaleosauridae и Pliosauridae - и ихтиозавр семья Офтальмозавры. Обнаружение нескольких останков молодых плезиозавров позволяет предположить, что они использовали богатые питательными веществами прибрежные воды в качестве защищенных мест для отела, а холод отпугивал хищников, таких как акулы. Большинство обнаруженных плезиозавров имели космополитическое распространение, тем не мение эндемичный там существовали формы, такие как Opallionectes и возможный новый вид криптоклидид. А сомнительный виды эласмозавридов Вулунгазавр, был назван в 1928 году одним из самых ранних описаний морских рептилий Австралии. Несколько моллюск, брюхоногие моллюски, аммонит, костлявая рыба, химера, и как кальмар белемнит останки также были восстановлены.[50] Прибрежный район, возможно, испытал зимнее замерзание, и эти рептилии, в ответ, мигрировали на север зимой, имели более активную метаболизм чем тропические рептилии, впали в спячку в пресноводных районах, как и в наши дни Американский аллигатор (Аллигатор миссисипиенсис), или были эндотермический похож на современный день кожистые морские черепахи (Dermochelys coriacea). Меньшее количество плезиозавров и большее количество ихтиозавров и морские черепахи в более северных областях Австралии указывает на предпочтение более холодных областей у плезиозавров.[50]

Несколько позднемеловых океанических плезиозавров и мозазавры были обнаружены в Новой Зеландии и Антарктиде, некоторые из них, например Мауизавр, будучи эндемичными, в то время как другие, такие как Прогнатодон, имеющий космополитическое распространение.[51][52] Эласмозавры и плиозавры известны от одного до трех видов из этой области. Открытие трех криптоклидид в Южном полушарии -Morturneria из Антарктиды, Aristonectes из Южной Америки и Kaiwhekea из Новой Зеландии - указывает на диверсификацию семейства в позднем меловом периоде этого региона и, возможно, на повышение продуктивности ранних Южный океан.[53]
Птерозавры

Две клады птерозавров представлены в раннемеловой Австралии, Pteranodontoidea и Ctenochasmatoidea, остается главным образом производным от Формация Тулебук и районы Квинсленда и Нового Южного Уэльса. Считается, что в Австралии мелового периода существовало как минимум шесть таксонов птерозавров, однако, учитывая фрагментарный характер останков, многие найденные окаменелости происходят от неустановленных птерозавров. Окаменелости были обнаружены на мелководье и в лагунах, что указывает на то, что они питались рыбой и другими водными обитателями. В ктенохазматиды были единственными археоптеродактилоиды дожить до мелового периода. Единственные остатки зуба птерозавра, обнаруженные в Австралии в раннем меловом периоде, принадлежат Mythunga и возможный позднемеловой период ангангуэрид. Mythunga По оценкам, размах крыльев составлял 4,5 метра (15 футов), что намного больше, чем у любого другого обнаруженного археоптеродактилоида, хотя возможно, что птерозавр больше связан с Anhangueridae или Орнитохейриды.[54][55][56] Тем не менее, птерозавры по-прежнему существуют в неполярных регионах Австралии, учитывая их способность перемещаться по воздуху, возможно, не нужно было пересекать сухопутный мост через полярные регионы, чтобы прибыть туда, то есть они никогда не населяли Южный полярный регион.[15][33]
Из позднемеловых птерозавров остались только останки семейства Azhdarchidae –Находится в Карнарвон и Перт бассейны в Западная Австралия –Были назначены таксон. Возможный представитель Ornithocheiridae был найден в позднемеловом периоде Западной Австралии, хотя ранее считалось, что это семейство вымерло в раннемеловом периоде.[54][55][56]
Млекопитающие

Семь млекопитающих были обнаружены в Австралии в раннем меловом периоде: неописанный орниторинхид, Криориктес, Колликодон, Аусктрибосфенос, Епископы, Стероподон, и Corriebaatar; все они были эндемичны для Австралии в то время. Вполне вероятно, что млекопитающие пересекли антарктический сухопутный мост между Австралией и Южной Америкой в раннем меловом периоде, и, вероятно, предки эндемичных млекопитающих Австралии прибыли в юрский период через Южную Америку. суперконтинент Пангея.[57][58]
Беспозвоночные
Несколько окаменелостей насекомые и ракообразные известны из южнополярных меловых отложений Новой Зеландии. Поздний мел Мангаотанский Монро Конгломерат находился на 68 ° ю.ш. и дал окаменелости Helastia sp.,[59] и краб Hemioon novozelandicum был найден в Swale алевролит, расположенная на 76 ° ю.ш. в конце Альбианский.[60] Несколько экземпляров насекомых также были обнаружены в Формация Тупуанги из Острова Чатем на широте 79 ° ю.ш. во время Сеноманский к Туронский.[61]
Смотрите также
- Бухта динозавров
- Динозавр мечтает
- Геология Антарктиды, включая палеонтологию
- Список австралийских и антарктических динозавров
- Тропические леса Гондваны
- Полярные леса мелового периода
Рекомендации
- ^ «Бухта динозавров, Австралия». Эта динамическая земля. USGS. Получено 14 марта 2015.
- ^ Vaughan, A.P.M .; Eagles, G .; Флауэрдью, М. Дж. (2012). «Свидетельства двухфазного события на Земле Палмера из пересекающихся структурных взаимосвязей и сроков внедрения интрузивной свиты Ласситер-Кост, Антарктический полуостров: последствия для конфигурации плит Южного океана в среднем меловом периоде» (PDF). Тектоника. 3 (1): TC1010. Bibcode:2012Tecto..31.1010V. Дои:10.1029 / 2011TC003006.
- ^ а б c Риффенбург 2007, п. 72.
- ^ Риффенбург 2007, п. 551.
- ^ Pirrie, D .; Crame, J. A .; Lomas, S.A .; Верховая езда, Дж. Б. (1997). «Позднемеловая стратиграфия района Адмиралтейского пролива, бассейн Джеймса Росс, Антарктида». Меловые исследования. 18 (1): 109–137. Дои:10.1006 / cres.1996.0052.
- ^ а б Weishampel, Дэвид Б. (2004). «Распространение динозавров (ранний мел, Австралазия)». В Weishampel, D. B .; Dodson, P .; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Калифорнийский университет Press. стр.573 –574. ISBN 978-0-520-24209-8.
- ^ Dettmann, M.E .; Molnar, R.E .; Дуглас, Дж. Г. (1992). «Австралийские меловые наземные фауны и флоры: биостратиграфические и биогеографические последствия». Меловые исследования. 13 (3): 207–262. Дои:10.1016/0195-6671(92)90001-7.
- ^ а б Хэй, В. В. (2008). «Развитие представлений о меловом климате и циркуляции океана». Меловые исследования. 29 (5): 725–753. Дои:10.1016 / j.cretres.2008.05.025.
- ^ Ladant, J .; Доннадье Ю. (2016). «Палеогеографическое регулирование ледниковых явлений в меловой надзеленой». Nature Communications. 7: 12771. Bibcode:2016НатКо ... 712771L. Дои:10.1038 / ncomms12771. ЧВК 5036002. PMID 27650167.
- ^ Beerling, D. J .; Осборн, К. П. (2002). «Физиологическая экология мезозойских полярных лесов в среде с высоким содержанием CO2». Анналы ботаники. 89 (3): 329–339. Дои:10.1093 / aob / mcf045. ЧВК 4233824. PMID 12096745.
- ^ Перальта-Медина, Э .; Фалькон-Лэнг, Х. Дж. (2012). «Состав и продуктивность меловых лесов по данным глобальной базы данных по ископаемой древесине». Геология. 40 (3): 219–222. Bibcode:2012Гео .... 40..219P. Дои:10.1130 / G32733.1.
- ^ Торн, В. (2001). «Сообщества растительности среднеюрского леса с высокой палеоширотой в Новой Зеландии». Палеогеография, палеоклиматология, палеоэкология. 168 (3–4): 273–289. Bibcode:2001ППП ... 168..273Т. Дои:10.1016 / S0031-0182 (01) 00203-6.
- ^ Поляк, М. (1999). «Строение леса приполярных широт из новозеландской юры». Палеогеография, палеоклиматология, палеоэкология. 147 (1–2): 121–139. Bibcode:1999ПП ... 147..121П. Дои:10.1016 / S0031-0182 (98) 00151-5.
- ^ Дуглас, Дж. Г .; Уильямс, Г. Э. (1982). «Южные полярные леса: раннемеловые флоры Виктории и их палеоклиматическое значение». Палеогеография, палеоклиматология, палеоэкология. 39 (3–4): 171–185. Bibcode:1982ППП .... 39..171Д. Дои:10.1016/0031-0182(82)90021-9.
- ^ а б c d е ж грамм Крэйм, Дж. А. (1992). «Позднемеловые палеосреды и биоты: перспектива Антарктики». Антарктическая наука. 4 (4): 371–382. Bibcode:1992AntSc ... 4..371C. Дои:10.1017 / S0954102092000555.
- ^ Pires, E. F .; Герра-Соммер М. (2011). «Анализ колец роста ископаемых хвойных лесов из раннего мела бассейна Арарипе (Бразилия)» (PDF). Anais da Academia Brasileira de Ciências. 83 (2): 409–423. Дои:10.1590 / S0001-37652011005000005. PMID 21625801.
- ^ а б Poole, I .; Gottwald, H .; Фрэнсис, Дж. Э. (2000). "Illicioxylon, стихия полярных лесов Гондваны? Позднемеловые и раннетретичные леса Антарктиды ». Анналы ботаники. 86 (2): 421–432. Дои:10.1006 / anbo.2000.1208.
- ^ а б c Викерс-Рич, П.; Рич, Т. Х. (1993). «Полярные динозавры Австралии». Scientific American. 269 (1): 50–55. Bibcode:1993SciAm.269a..50V. Дои:10.1038 / scientificamerican0793-50. JSTOR 24941547.
- ^ а б Готвальд, Х. (2001). "Monimiaceae sensu lato, элемент полярных лесов Гондваны: свидетельства позднемеловой - раннетретичной древесной флоры Антарктиды ». Австралийская систематическая ботаника. 14 (2): 207–230. Дои:10.1071 / SB00022.
- ^ Константин, А .; Чинсамы-Туран, А .; Rich, P. V .; Рич, Т. (1998). «Перигляциальные среды и полярные динозавры». Южноафриканский научный журнал. 94 (3): 137–141.
- ^ Close, R. A .; Vicker-Rich, P .; Trusler, P .; Chiappe, L.M .; О'Коннор, Джингмай; Rich, T. H .; Kool, L .; Комаровер, П. (2008). «Самая ранняя гондванская птица из мелового периода юго-востока Австралии». Журнал палеонтологии позвоночных. 29 (2): 616–619. Дои:10.1671/039.029.0214.
- ^ Аллея, Н. Ф .; Фрейкс, Л. А. (2003). «Первое известное меловое оледенение: пачка тиллитов Ливингстона формации Каднауи, Южная Австралия». Австралийский журнал наук о Земле. 50 (2): 139–144. Bibcode:2003AuJES..50..139A. Дои:10.1046 / j.1440-0952.2003.00984.x.
- ^ а б Кир, Б. П. (2004). «Останки плезиозавра из меловых высокоширотных неморских отложений на юго-востоке Австралии». Журнал палеонтологии позвоночных. 26 (1): 196–199. Дои:10.1671 / 0272-4634 (2006) 26 [196: PRFCHN] 2.0.CO; 2.
- ^ Прайс, Г. Д. (1999). «Свидетельства и последствия полярных льдов во время мезозоя». Обзоры наук о Земле. 48 (3): 183–210. Bibcode:1999ЕСРв ... 48..183П. Дои:10.1016 / S0012-8252 (99) 00048-3.
- ^ Falcon-Lang, H.J .; Кантрилл, Д. Дж. (2001). «Фенология листьев некоторых полярных лесов среднего мелового периода, остров Александр, Антарктида». Геологический журнал. 138 (1): 39–52. Bibcode:2001ГеоМ..138 ... 39F. Дои:10.1017 / S0016756801004927.
- ^ а б Риффенбург 2007, п. 413.
- ^ Dettmann, M.E .; Ярзен, Д. М. (1991). «Пыльцевые свидетельства позднемеловой дифференциации Proteaceae в южных полярных лесах». Канадский журнал ботаники. 69 (4): 901–906. Дои:10.1139 / b91-116.
- ^ Francis, J. E .; Пул, И. (2002). «Меловой и раннетретичный климат Антарктиды: данные по ископаемой древесине». Палеогеография, палеоклиматология, палеоэкология. 182 (1–2): 47–64. Bibcode:2002ППП ... 182 ... 47Ф. Дои:10.1016 / S0031-0182 (01) 00452-7.
- ^ Замек горка комплекс в Окаменелости.org
- ^ Комплекс Скуа Бэй в Окаменелости.org
- ^ Половина трехточечной сборки в Окаменелости.org
- ^ Формация Собрал в Окаменелости.org
- ^ а б c d е ж Мольнар Р. Э. (1989). «Наземные четвероногие в Антарктиде мелового периода». В Крейне, Дж. А. (ред.). Происхождение и эволюция антарктической биоты. Геологическое общество, Лондон, Специальные публикации. 47. Специальные публикации Геологического общества. С. 131–140. Bibcode:1989ГСЛСП..47..131М. CiteSeerX 10.1.1.1010.6989. Дои:10.1144 / GSL.SP.1989.047.01.11.
- ^ Риффенбург 2007, п. 146.
- ^ а б c d Tambussi, C .; Оспиталече, К. А. (2007). «Антарктические птицы (Neornithes) в меловое – эоценовое время» (PDF). Revista de la Asociación Geológica. 62 (4): 604–617.
- ^ Hackett, S.J .; Kimball, R.T .; Reddy, S .; Боуи, Р. К. К .; Braun, E.L .; Браун, М. Дж. (2008). «Филогеномное исследование птиц раскрывает их эволюционную историю». Наука. 320 (5884): 1763–1768. Bibcode:2008Sci ... 320.1763H. Дои:10.1126 / science.1157704. PMID 18583609.
- ^ а б Benson, R. B.J .; Rich, T. H .; Vickers-Rich, P .; Холл, М. (2012). «Фауна теропод из Южной Австралии указывает на высокое полярное разнообразие и обусловленную климатом провинциальность динозавров». PLOS One. 7 (5): e37122. Bibcode:2012PLoSO ... 737122B. Дои:10.1371 / journal.pone.0037122. ЧВК 3353904. PMID 22615916.
- ^ а б Rich, T. H .; Vickers-Rich, P .; Ганглофф, Р. А. (2002). «Полярные динозавры» (PDF). Наука. 295 (5557): 979–980. Дои:10.1126 / science.1068920. PMID 11834803.
- ^ а б c d Poropat, S. F .; Mannion, P.D .; Upchurch, P .; Hocknull, S.A .; Кир, Б. П. (2016). «Новые австралийские зауроподы пролили свет на палеобиогеографию динозавров мелового периода». Научные отчеты. 6 (34467): 34467. Bibcode:2016НатСР ... 634467П. Дои:10.1038 / srep34467. ЧВК 5072287. PMID 27763598.
- ^ Case, J. A .; Martin, J. E .; Регуэро, М. (2007). «Дромеозавр из маастрихта острова Джеймса Росса и позднемеловой антарктической фауны динозавров». Геологическая служба США и национальные академии. CiteSeerX 10.1.1.546.3890.
- ^ а б Richards, T.HV .; Richards, T. H .; Викерс-Рич, П. (2000). "Откуда мы знаем; куда мы идем?". Динозавры тьмы. Издательство Индианского университета. ISBN 978-0-253-33773-3.
- ^ Bell, P.R .; Снайвели, Э. (2007). «Парад полярных динозавров: обзор миграции динозавров». Алчеринга: Австралазийский журнал палеонтологии. 32 (3): 271–284. Дои:10.1080/03115510802096101.
- ^ Chinsamy, A .; Rich, T .; Викерс-Рич, П. (1998). «Гистология костей полярного динозавра». Журнал палеонтологии позвоночных. 18 (2): 385–390. Дои:10.1080/02724634.1998.10011066.
- ^ а б Dettmann, M.E .; Molnar, R.E .; Дуглас, Дж. Г .; Burger, D .; Филдинг, К. (1992). «Австралийские меловые наземные фауны и флоры: биостратиграфические и биогеографические последствия». Меловые исследования. 13 (3): 207–262. Дои:10.1016/0195-6671(92)90001-7.
- ^ Woodward, H.N .; Rich, T. H .; Chinsamy, A .; Викерс-Рич, П. (2011). «Динамика роста полярных динозавров Австралии». PLOS One. 6 (8): e23339. Bibcode:2011PLoSO ... 623339W. Дои:10.1371 / journal.pone.0023339. ЧВК 3149654. PMID 21826250.
- ^ Мартин, А. Дж. (2009). «Норы динозавров в группе Отуэй (Альб) в Виктории, Австралия, и их связь с полярными средами мелового периода». Меловые исследования. 30 (5): 1223–1237. Дои:10.1016 / j.cretres.2009.06.003.
- ^ Уайт, М. А .; Bell, P.R .; Cook, A. G .; Poropat, S. F .; Эллиотт, Д. А. (2015). "Зубной камень Австраловенатор Винтоненсис (Theropoda, Megaraptoridae); последствия для зубного ряда мегарапторид ". PeerJ. 3: e1512. Дои:10.7717 / peerj.1512. ЧВК 4690360. PMID 26713256.
- ^ Molnar, R.E .; Wiffen, J .; Хейс, Б. (1998). «Вероятная кость теропод из новейшей юры Новой Зеландии». Новозеландский журнал геологии и геофизики. 41 (2): 145–148. Дои:10.1080/00288306.1998.9514798.
- ^ а б c Уоррен, А .; Kool, L .; Cleeland, M .; Викерс-Рич, П. (1991). "Раннемеловой лабиринтодонт". Алчеринга. 15 (3–4): 327–332. Дои:10.1080/03115519108619027.
- ^ а б c Кир, Б. П. (2006). «Морские рептилии из нижнего мела Южной Австралии: элементы сообщества высоких широт и холодной воды». Палеонтология. 49 (4): 837–856. Дои:10.1111 / j.1475-4983.2006.00569.x.
- ^ Martin, J. E .; Фернандес, М. (2007). "Синонимия позднемелового рода мозазавров (Squamata)" Лакумазавр из Антарктиды с Танивазавр из Новой Зеландии и его влияние на сходство фауны в провинции Уэдделлиан ". Геологический журнал. 42 (2): 203–211. Дои:10.1002 / gj.1066.
- ^ Виффен, Дж. (1989). «Новые мозазавры (Reptilia; семейство Mosasauridae) из верхнего мела Северного острова, Новая Зеландия». Новозеландский журнал геологии и геофизики. 33 (1): 67–85. Дои:10.1080/00288306.1990.10427574.
- ^ Cruickshank, A.R. I .; Фордайс, Р. В. (2003). «Новая морская рептилия (Sauropterygia) из Новой Зеландии: еще одно свидетельство австральной радиации криптоклидидных плезиозавров в позднем меловом периоде». Палеонтология. 45 (3): 557–575. Дои:10.1111/1475-4983.00249.
- ^ а б Broughman, T .; Smith, E.T .; Белл, П. Р. (2017). «Изолированные зубы Anhangueria (Pterosauria: Pterodactyloidea) из нижнего мела Лайтнинг-Ридж, Новый Южный Уэльс, Австралия». PeerJ. 3 (5): e3256. Дои:10.7717 / peerj.3256. ЧВК 5419211. PMID 28480142.
- ^ а б Keller, A. W. A .; Rich, T. H .; Costa, F. R .; Vickers-Rich, P .; Kear, B.P .; Уолтерс, М. (2009). «Новые изолированные птеродактилоидные кости из формации Альбиан Тулебук (западный Квинсленд, Австралия) с комментариями об австралийской фауне птерозавров». Алчеринга. 34 (3): 219–230. Дои:10.1080/03115511003656552.
- ^ а б Kear, B.P .; Дьякон, Г. Л .; Сиверсон, М. (2009). «Остатки позднемелового птерозавра из Молекап-Гринсанд в Западной Австралии». Алчеринга. 34 (3): 273–279. Дои:10.1080/03115511003661651.
- ^ Rich, T. H .; Vickers-Rich, P .; Фланнери, Т. Ф .; Кир, Б. П. (2009). «Австралийский мультитуберкулез и его палеобиогеографические последствия». Acta Palaeontologica Polonica. 54 (1): 1–6. Дои:10.4202 / app.2009.0101.
- ^ Арчер, М .; Фланнери, Т. Ф .; Ritchie, A .; Мольнар, Р. Э. (1985). «Первое мезозойское млекопитающее из Австралии - монотремы раннего мела». Природа. 318 (6044): 363–366. Bibcode:1985Натура.318..363А. Дои:10.1038 / 318363a0.
- ^ Монро Конгломерат в Окаменелости.org
- ^ Swale Stream в Окаменелости.org
- ^ Залив Вайхере, остров Питт в Окаменелости.org
дальнейшее чтение
- Риффенбург, Б. (2007). Энциклопедия Антарктики. 1. Тейлор и Фрэнсис. ISBN 978-0-415-97024-2.
- Dutra, T. L .; Баттен, Д. Дж. (2000). «Флоры верхнего мела острова Кинг-Джордж в Западной Антарктиде и их палеоэкологические и фитогеографические значения». Меловые исследования. 21 (2–3): 181–209. Дои:10.1006 / cres.2000.0221.
- Филкорн, Х. Ф. (1994). «Ископаемые кораллы-склерактинии из бассейна Джеймса Росс, Антарктида». Серия антарктических исследований. 65: 1–96.
- Harris, A.C .; Рейн, Дж. (2002). «Склерит из позднемелового мотылька (Insecta: Lepidoptera) из ущелья Ракайя, Кентербери, Новая Зеландия». Журнал Королевского общества Новой Зеландии. 32 (3): 457–462. Дои:10.1080/03014223.2002.9517704.
- Poole, I .; Mennega, A.MW .; Кантрилл, Д.Дж. (2003). «Вальдивские экосистемы в позднем меловом и раннем третичном периоде Антарктиды: дальнейшие свидетельства из ископаемой древесины миртовых и эвкрифических пород». Обзор палеоботаники и палинологии. 124 (1–2): 9–27. Дои:10.1016 / S0034-6667 (02) 00244-0. HDL:1874/31608.
- Рич, Т. Х. В. (2007). Полярные динозавры Австралии. Музей Victoria Nature Series. Музей Виктории. ISBN 978-0-9758370-2-3.
- Спеден, И. (1973). «Распространение, стратиграфия и стратиграфические отношения меловых отложений, западная часть полуострова Раукумара, Новая Зеландия». Новозеландский журнал геологии и геофизики. 16 (2): 243–268. Дои:10.1080/00288306.1973.10431456.
- Stilwell, J.D .; Vitacca, J.J .; Мэйс, К. (2016). «Южнополярные тепличные насекомые (Arthropoda: Insecta: Coleoptera) из формации Тупуанги среднего мела, острова Чатем, восточная Зеландия». Алчеринга. 40 (4): 502–508. Дои:10.1080/03115518.2016.1144385.