Катопсбаатар - Catopsbaatar
| Катопсбаатар | |
|---|---|
 | |
| Наиболее полный скелет (образец PM120 / 107) показан сверху (слева, A) и снизу (справа, B) в сохраненном виде, со схемами, показывающими отдельные кости | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Класс: | Млекопитающие |
| Заказ: | †Мультитуберкулята |
| Семья: | †Djadochtatheriidae |
| Род: | †Катопсбаатар Kielan-Jaworowska, 1994 |
| Разновидность: | †С. catopsaloides |
| Биномиальное имя | |
| †Катопсбаатар катопсалоидес (Келан-Яворовская, 1974) | |
| Синонимы | |
| |
Катопсбаатар это род из многотуберкулезный, вымерший порядок из грызун -подобно млекопитающие. Он жил в том, что сейчас Монголия в конце Кампанский возраст Поздний мел эпоха около 72 миллионов лет назад. Первые окаменелости были собраны в начале 1970-х годов, и животное было названо новым видом этого рода. Джадохтатерий в 1974 г., D. catopsaloides. В конкретное имя относится к сходству животного с роду Катопсалис. Вид переведен в род Катопсалис в 1979 г. и получил свой род (Катопсбаатар, Греческое и монгольское для «видимого героя») в 1994 г. Известно пять черепов, один коренной зуб и один скелет с черепом; последний - наиболее полный экземпляр рода. Катопсбаатар был членом семьи Djadochtatheriidae.
Череп Катопсбаатар был до 70 мм (2,8 дюйма) в длину и, как и при других мульти-туберкулезных заболеваниях, был пропорционально большим. По внешнему виду головы этих животных могли быть похожи на головы грызунов. Череп был массивным и широким, с скуловые дуги сильно расширен в стороны. Глазницы были меньше и располагались дальше назад, чем у его родственников, а морда была более вытянутой. Катопсбаатар имел полукруглые гребни сбоку черепа, к которым прикреплялись мышцы челюсти. В нижняя челюсть был крепким и очень вытянутым. У него был очень прочный резцы, и щечные зубы с множественными куспиды (в честь чего названы мультитуберкуляты). В кости таза отличается от таковых других мультитуберкулезных групп тем, что они не слились друг с другом. Катопсбаатар имел шпоры на щиколотках, как у мужчин утконос и ехидна, без доказательств яд канал (присутствует в бывшем).
Шпоры Катопсбаатар и другие Мезозойский млекопитающие могли использоваться для защиты от теропод динозавры и другие хищники. Считается, что мультитуберкулезные заболевания дали живорождение, и тот факт, что у них были волосы, указывает на то, что они теплокровный («теплокровный»). Мультитуберкуляты были бы всеядный; Катопсбаатар имел мощные мускулы челюсти, а его резцы были хорошо приспособлены для грызения твердых семян с помощью жевательного движения назад. Считается, что у мультитуберкулятов была распростёртая осанка и Катопсбаатар мог прыгать. Катопсбаатар известно из Формация Барун Гойот, которому, как полагают, около 72 миллионов лет.
Таксономия

В 1970 и 1971 годах Польско-монгольские палеонтологические экспедиции собрали окаменелости млекопитающих из Формация Барун Гойот на Красных ложе Хермийн Цав (также пишется "Хермин Цав") в районе Монголии пустыня Гоби. Около 100 экземпляров, извлеченных из четырех местонахождений, размещены на Польская Академия Наук в Варшаве. Две трети собранных образцов были многотуберкулезные: вымерший порядок млекопитающих с грызун -подобный зубной ряд, названный в честь многочисленных куспиды (или же бугорки ) на их коренные зубы. В 1974 г. польский палеонтолог. Зофья Киелан-Яворовская назвал новый вид монгольского многотуберкулезного рода Джадохтатерий так как D. catopsaloides, с образцом ZPAL MgM − I / 78 из польской коллекции в качестве голотип. В конкретное имя относится к сходству животного с североамериканскими видами Катопсалис Джойнери, которого Келан-Яворовская считала возможным потомком. Экземпляр, собранный в местонахождении Эрмиин Цав I, представляет собой почти полный череп подростка с поврежденными частями черепа. Киелан-Яворовска также отнесла к этому виду другие образцы: поврежденный череп без нижней челюсти (ZPAL MgM − I / 79, взрослый), череп с частичной нижней челюстью (ZPAL MgM − I / 80) и коренной зуб с фрагментом. челюсти (ZPAL MgM − I / 159 из Хулсана, единственный экземпляр не из местонахождений Эрмиин-Цав I и II).[1][2][3][4]

Килан-Яворовска и американский палеонтолог Роберт Э. Слоан считали род Джадохтатерий а младший синоним из Катопсалис, и создал новая комбинация С. catopsaloides в 1979 г.[5] Американские палеонтологи Нэнси Б. Симмонс и Мяо Десуи провели исследование 1986 г. кладистический анализ что указывает на то, что Катопсалис был парафилетический таксон (неестественная группировка видов) и С. catopsaloides требуется собственное родовое имя.[6] Киелан-Яворовска последовала предложению Симмонса и Мяо, двигаясь С. catopsaloides к своему собственному монотипный род в 1994 г., Катопсбаатар. Слово Catops происходит из Греческий Катоптос («видимый» или «очевидный»); баатар на монгольском языке означает «герой», и это имя относится к Катопсбаатарs сходство с родом Катопсалис (как и в случае с конкретным именем).[7] Название Катопсалис сам состоит из греческих слов, обозначающих «видимые» и «ножницы для резки» (псалис).[8][7] Слово баатар используется как суффикс в названиях многих многотуберкулезных родов и намекает на столицу Монголии Улан-Батор, что само по себе означает «красный герой».[9]
Позже, в 1994 году, Киелан-Яворовска и российский палеонтолог Петр П. Гамбарян упомянули хвостовые (хвостовые) позвонки, которые, возможно, принадлежали Катопсбаатар; эта атрибуция сомнительна, так как они могут вместо этого принадлежать Томбаатар (назван в 1997 г.).[10][11] Четвертый череп (ПИН 4537/4, молодой), обнаруженный во время Советско-Монгольской экспедиции 1975 года, упоминался Гамбарян и Киелан-Яворовской в 1995 году.[12] Канадский палеонтолог Филип Дж. Карри нашел новый Катопсбаатар Образец во время экспедиции динозавров в Гоби в 1999 году, организованной американской компанией Nomadic Expeditions Company. Размещен в Монгольская Академия Наук в Улан-Баторе, как PM120 / 107, этот наиболее полностью сохранившийся известный экземпляр состоит из полного черепа (который может быть слегка приплюснут) и частичного скелета взрослого человека. У особи довольно полные передние и задние конечности, которые до этого не были известны для этого рода и которые, как правило, редко сохраняются у мультитуберкулятов. это тазовый ilia были украдены и уничтожены школьником на гастролях в Музей естественной истории Осло, где он готовился в 2000 году. Образец был обнаружен в 2002 году Киелан-Яворовской, норвежским палеонтологом. Йорн Хурум, Карри и монгольский палеонтолог Ринчен Барсболд, который также упомянул еще один череп (PIN 4537/5, молодой), найденный во время экспедиции 1975 г.[11][13][3] Catopsalis joyneri, основа имени С. catopsaloides, был переведен в новый род Валенопсалис в 2015 году.[8][1]
Эволюция
Катопсбаатар принадлежал к отряду Multituberculata, группе внутри Аллотерия (инфракласс млекопитающие за пределами Терия, группа, содержащая современные плаценты и сумчатые ). Мультитуберкулятам свойственны премоляры и моляры с множеством низких бугорков, расположенных продольными рядами. Мультитуберкуляты - самая известная группа млекопитающих из Мезозойская эра, когда динозавры преобладают; хотя самые ранние остатки мультитуберкулеза относятся к Юрский период, группа известна еще недавно как Эоценовая эпоха (тем самым выживая Меловое – палеогеновое вымирание ). Группа могла вымереть из-за конкуренции с евтерский млекопитающие, такие как грызуны. Мультитуберкулезные инфекции были известны в основном по зубам и челюстям до 1920-х годов, когда были обнаружены более полные образцы - сначала в Азии, а затем в других местах.[4] Посткраниальный кости (остальная часть скелета, кроме черепа) остаются редкими.[13]
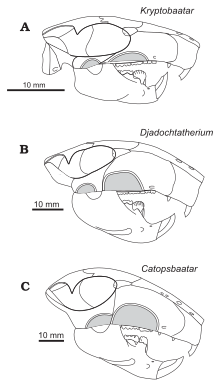
Келан-Яворовская первоначально засекречена Катопсбаатар как член многотуберкулезной семьи Taeniolabididae в 1974 г.[1] В 1994 году она предложила Джадохтатерий был близок к Катопсбаатарс происхождение.[7] Она и Хурум назвали новую семью мультитуберкулезных больных, Djadochtatheriidae (который они поместили в новый подотряд Джадохтатерия ), в 1997 г. В семью входили роды Джадохтатерий, Катопсбаатар, Криптобатор, и Томбаатар, все из пустыни Гоби. Семейство отличается от других мультитуберкулезных (и других млекопитающих) тем, что передние края его морды сливались с скуловые дуги (скулы), придавая морде трапеция форма при взгляде сверху. В общем, у других млекопитающих есть морды, где боковые края изогнуты внутрь перед скуловыми дугами.[14] Kielan-Jaworowska и Hurum пересмотрели высшие ранги в Multituberculata в 2001 году, заменив подотряд Djadochtatheria надсемейством Djadochtatherioidea (помещенный в подотряд Чимолодонта ).[15]
Следующее кладограмма показывает размещение Катопсбаатар среди других мультитуберкулезных заболеваний по данным Kielan-Jaworowska and Hurum, 1997:[14]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Описание
Череп

Самый полный взрослый Катопсбаатар Череп (образец PM 120/107) имеет длину 63 мм (2,5 дюйма) и ширину 55 мм (2,2 дюйма) с длиной нижней челюсти 41 мм (1,6 дюйма). Для сравнения: череп молодого голотипа (ZPAL MgM-I / 78) имеет длину около 53 мм (2,1 дюйма) и ширину 56 мм (2,2 дюйма) с длиной нижней челюсти 35 мм (1,4 дюйма). Самый большой череп взрослого человека (ZPAL MgM-I / 79) имеет длину 70 мм (2,8 дюйма), но, поскольку он неполный, его другие размеры неизвестны. Катопсбаатар был крупнее своих сородичей, Криптобатор и Джадохтатерий.[3] Мультитуберкулы имели относительно большие черепа и короткие шеи; их черепа были пропорционально длиннее и шире, чем у грызунов и сумчатых того же размера.[4] По внешнему виду их головы могли быть похожи на головы грызунов.[12]
Череп Катопсбаатар был массивным, с широким полем спереди. По средней линии он был короче, чем по бокам, потому что затылочный гребень задняя часть головы изогнута внутрь посередине, образуя выемку на заднем крае черепа, если смотреть сверху. Скуловые дуги были сильно расширены в стороны, ширина черепа (поперек дуг) составляла около 85 процентов от длины черепа. Передняя часть орбиты (глазницы) располагались дальше назад, чем у других джадохтатериид, что привело к более удлиненной морде (65% длины черепа) и небольшим орбитам. В предчелюстная кость (передняя кость верхней челюсти) вытянута менее чем на две трети длины морды перед глазами, короче, чем у Криптобатор. Предчелюстная кость состоит из частей лица и неба; djadochtatheriids имел предчелюстной гребень на границе между ними (виден при осмотре черепа снизу). Небная часть предчелюстной кости была вогнутой, с множеством беспорядочно расположенных питательных веществ. отверстие (проемы). В носовая кость - образующий верхнюю часть рыла - был относительно широким (становился шире к спине), а его передняя часть была покрыта неравномерно расположенными сосудистыми отверстиями. В шов между носовой и лобные кости был менее направлен вперед посередине, чем Катопсбаатарродственники.[3]

В верхняя челюсть (основная часть верхней челюсти) была обширной и составляла большую часть стороны морды. В нем были все верхние зубы, кроме резцы. В подглазничное отверстие (отверстия в нижней части верхней челюсти) у одних экземпляров были щелевидными, у других закругленными, их количество варьировалось от одного до трех. Одна из самых характерных черт лица Катопсбаатар был очень большой передний скуловой гребень по бокам верхней челюсти (место прикрепления челюстных мышц). Он был намного выше, чем у других джадохтатериид, за исключением Джадохтатерий, от которого он отличался тем, что гребень был полукруглым, а не примерно трапециевидным (у других родов гребни эллиптические). Передний край переднего скулового гребня был утолщен, что приводило к выпуклости на боковом крае рыла при осмотре сбоку и сверху. Нижняя часть шва между верхней челюстью и чешуйчатая кость проходит по задней границе переднего скулового валика. Небные отростки верхней челюсти составляют большую часть неба. В большое небное отверстие имел неглубокие бороздки, идущие вперед. В постпелатиновый тор (костный выступ на небе) Катопсбаатар был менее заметным, чем у Томбаатар. Лобная кость была большой и составляла большую часть крыши черепа. Шов между лобной и теменные кости образует U-образную часть в середине, которая направлена назад - аналогично условию в Криптобатор но менее глубокие, с более мелкими U-образными структурами по бокам. Это отличалось от Джадохтатерийс более узким V-образным швом между лобной и теменной костями.[3]

Промежуточный скуловой гребень на чешуйчатой кости (также для прикрепления челюстных мышц) был намного меньше и ниже, чем передний скуловой гребень перед ним. Катопсбаатар отличается от других джадохтатериид тем, что промежуточный гребень контактировал с передним гребнем своим передним краем. Задний скуловой гребень в нижней части чешуйчатой кости был самым слабым из трех гребней и отмечен лишь вдавлением. В заглазничный отростки за орбитой на теменной кости были очень длинными, а теменные гребни тянулись от заднего края заглазничных отростков навстречу друг другу (но не доходили). Затылочный гребень был очень заметным и расширялся в стороны, образуя «крылья». В затылочный пластина была почти вертикальной и слегка вогнутой и при осмотре черепа сверху закрывалась затылочным гребнем. Предполагается, что затылочная пластинка сохранилась не полностью, но отошла назад и вверх от позвоночника. затылочные мыщелки (как у некоторых родственников).[3] Орбитовисочная сосудистая система внутри черепа Катопсбаатар мало чем отличались от родственных родов.[16]
Катопсбаатарс нижняя челюсть был прочным и очень удлиненным. В диастема (промежуток между передними и щечными зубами) был вогнутым и увеличивался на 20 процентов зубная кость (основная кость нижней челюсти). При взгляде сверху диастема образовывала широкую полку, наклоненную вниз на внутренней стороне челюсти. Маленький подбородочное отверстие был близко к верхнему среднему краю диастемы. В венечный отросток нижней челюсти по-видимому, был относительно длиннее и уже, чем у других джадохтатериоидов. Он был отделен от альвеолярный отросток (где находятся зубья) широкой бороздкой. В мыщелок нижней челюсти (который сочленяется с черепом) был немного выше уровня коренных зубов. Передняя часть массеретического гребня была очень выступающей, образуя выпуклость, известную как массеретический выступ. Прочность этого гребня и наличие выпуклости варьируются среди родственных родов. Жевательный ямка (ямка) перед жевательной ямкой, вероятно, была более отчетливой, чем у других джадохтатериоидов. Каждая половина нижнечелюстной симфиз (где две половинки нижней челюсти соединяются) имела форму перевернутой слезы. В крыловидная ямка внутренняя сторона нижней челюсти была очень большой и занимала большую часть задней части зубной кости. Нижняя часть этой ямки имела границу, известную как крыловидная полка.[3]
Зубы

В зубная формула (количество зубов каждого типа в зубном ряду млекопитающего) Катопсбаатар был 2.0.3.21.0.2.2 (два резца, нет клыки, три премоляра и два моляра в половине верхнего зубного ряда и один резец, без клыков, два премоляра и два моляра в половине нижнего). Для сравнения: зубная формула человека 2.1.2.2–32.1.2.2–3. Каждый зуб у млекопитающего обозначается буквой и номером по положению (I для резца, C для клыка, P для премоляра, M для моляра); буквы для зубов верхней челюсти пишутся с заглавной буквы, но не для зубов нижней. Формула бугров показывает расположение и количество бугорков в последовательных рядах зуба, от внешней к внутренней стороне; каждая строка отделяется двоеточием.[3][4][11]
Цимолодонт, Катопсбаатар резцов I1 не было.[14] Передние верхние резцы I2 Катопсбаатар были очень крепкими и имели резко ограниченный диапазон эмаль. Два резца слегка сходились к середине, касаясь друг друга. Меньший резец I3 сзади имел конусовидную форму. В альвеола (зубная лунка) из Катопсбаатарs Резец I3 образовался на уровне предчелюстной кости, а не на уровне предчелюстной кости и верхней челюсти (в отличие от Томбаатар). Передние верхние премоляры P1 и P3 присутствовали только у молодых особей (лиственный ), исчезающие (вместе со своими альвеолами) у пожилых людей. P1, по-видимому, имел две бугорки, был однокорневым и имел конусообразную тупую форму. Корона. P3 был однокорневым и меньше P1. Формула бугорка премоляра P4 была 5-4: 1, центральный бугорок был самым большим. P4 из Катопсбаатар был почти трапециевидной формы (в отличие от Джадохтатерий и Криптобатор, где она имеет форму полумесяца), меньшего размера и без гребней. Катопсбаатар также отличается наличием только трех верхних премоляров без P2 (общая черта с Томбаатар). У других млекопитающих потеря зубов обычно происходит в начале или в конце зубного ряда, а не в середине (как у мультитуберкулезных). Формула бугорка моляра M1 была 5-6: 5-6: 4 с внутренним гребнем, простирающимся примерно на 75 процентов длины зуба. Бугорки M1 были острыми и неизношенными у молодых, но изношенными и вогнутыми у более старых животных. Формула куспида моляра М2 была 2: 2−3: 2−3.[3][4]
Катопсбаатар имел единственную нижнюю пару резцов, характерную для мультитуберкулезных, которые были очень крепкими и сжатыми вбок. У него была резко ограниченная полоса эмали, и она постоянно росла. Премоляр p3 был очень маленьким и полностью прилегал к нижней диастеме под большим p4. Лопаткообразный p4 имел примерно трапециевидную форму на виде сбоку и имел три бугорка вдоль горизонтального верхнего края и один бугорок на внешней задней стороне. Р4 не имел выступов на внешней и внутренней стороне, как у других мультитуберкулезных больных. Моляр m1 был почти симметричным, а его формула бугров составляла 4: 4, при этом размер бугорков уменьшался по направлению к спине. M2 имел формулу заострения 2−3: 2, большинство экземпляров 2: 2. Бугорки с внутренней стороны шире, чем с внешней, внутренний ряд бугорков короче внешнего, задний край зуба расположен наклонно.[3]
Посткраниальный скелет

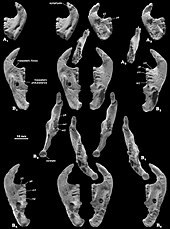
Единственный Катопсбаатар Образец, сохраняющий посткраниальный скелет, - PM120 / 107, фрагментарный. Он включает элементы, которые неизвестны или сохранились не полностью у других мульти-туберкулезных заболеваний. Один поясничный позвонок (пятый или шестой, между грудной клеткой и тазом) имел остистый отросток, который был толстым при виде сбоку и длинным при взгляде сверху. В ключица был немного менее изогнутым, чем у Криптобатор (напоминающий изогнутый стержень, расширяющийся на каждом конце), и его размер составляет около 24,8 мм (0,98 дюйма). Верхняя часть скапулокоракоид был относительно широким при виде сбоку, а нижняя часть была очень узкой. Когда он был готов, он, вероятно, был около 60 мм (2,4 дюйма) в длину. Сохранившаяся часть плечевая кость (кость плеча) была около 37,5 мм (1,48 дюйма) в длину. Его древко было треугольным в поперечном сечении, относительно узким, если смотреть сверху, и большую часть его ширины занимала межбубная борозда. Эта борозда была ограничена сбоку гребнем гребня. большой бугорок, средняя часть которого образовывала дельтовидно-грудной гребень. Локтевой мыщелок, где локтевая кость нижней части руки, сочлененной с плечевой костью, был более выраженным, чем лучевой мыщелок (где сочленяется лучевая кость), овальной формы и отделен от лучевого мыщелка канавкой. Радиус был около 26 мм (1,0 дюйма) в длину с выступающей головкой. Его древко было гладким, сжатым сверху вниз и овальным в поперечном сечении. Локтевая кость была сдавлена вбок, более плоская, чем лучевая, и сохранилась примерно на 40 мм (1,6 дюйма) в длину.[13]

Контакт из седалищная кость к подвздошной кости и лобок таза не срослось, а передний конец седалищной кости образовал морщинистый шов. Лобок был примерно треугольной формы, с грубым швом для подвздошной кости вверху и глубокой бороздкой для седалищной кости внизу спереди. Кости таза у образца PM120 / 107 отличались от таковых у других мультитуберкулезных больных тем, что не срослись вместе. Наличие швов в тазу PM120 / 107 указывает на то, что это был несовершеннолетний, хотя череп выглядит взрослым; смысл этого расхождения неизвестен. Бедренная кость (бедренная кость) была пропорционально аналогична кости бедра. Евкосмодон и Nemegtbaatar- меньше первого, но больше второго. Бедренная кость была толстой относительно своей длины и могла быть около 56 мм (2,2 дюйма) в длину. В большеберцовая кость голени была около 35,8 мм (1,41 дюйма) в длину. При осмотре сзади на верхней стороне большеберцовая кость имеет глубокую выемку (полость), что может быть характерно для мультитуберкулеза.[13]
В отличие от большинства других мультитуберкулезных и других млекопитающих, пяточная кость на тыльной стороне стопы имелся короткий клубень пяточной кости (как у некоторых кенгуру ), с расширенным проксимальным отростком в форме наковальни, сильно загнутым вниз и в стороны. Катопсбаатар на внутренней стороне лодыжки имелась костная костная ткань os calcaris, что также наблюдается у современных мужчин монотремы (в утконос и ехидна ) и других мезозойских млекопитающих. Кость os calcaris имела пластинчатую форму прямоугольной формы; как и в случае с монотремами, он поддерживал кальциевый рог в формировании шпора на внешней стороне предплюсна (скопление костей стопы). В отличие от других мезозойских млекопитающих, эти два элемента не были слиты вместе в мультитуберкулятах. Cornu calcaris имел треугольную форму с вогнутостью посередине и имел длину 13 мм (0,51 дюйма). Шпора была уплощена и была толще в месте соединения с os calcaris (где они соединялись несколькими гребнями). Поскольку шпора PM120 / 107 могла быть смещена из своего исходного положения, неизвестно, обращена ли она внутрь (как у утконоса). В отличие от утконоса, не было никакого впечатления от канала для яд. Cornu calcaris из Катопсбаатар был окостеневший (превратился в кость) и был бы покрыт кератин (роговой покров на ногтях и копытах). Cornu calcaris утконоса состоит только из кератина и является полым.[13][17]
Палеобиология
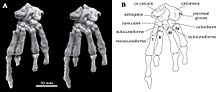
Hurum, Zhe-Xi Luo и Kielan-Jaworowska предположили в 2006 году, что шпоры на лодыжках мезозойских млекопитающих (таких как Катопсбаатар) мы гомологичный с монотремами, и были базальный (или «примитивный») признак, утраченный более поздними терианскими млекопитающими. Самец утконоса использует шпору, чтобы доставить яд из железы, но неизвестно, были ли вымершие группы ядовитыми. Мезозойские млекопитающие были в основном небольшими (за исключением таких, как лиса размером с лису). Репеномам ) и, хотя они были слишком малы, чтобы стать добычей для крупных теропод ими могли питаться динозавры, более мелкие тероподы, большие ящерицы, крокодилы и птицы. Например, челюсти млекопитающих были обнаружены в брюшной полости особи маленького теропод. Синозауроптерикс; челюсти принадлежали Жангеотерий, который также имел шпоры, и многотуберкулезный Синобатор. Поскольку в мезозое на Земле доминировали динозавры, этот период получил название "Темные времена "истории млекопитающих. Шпора, которая была бы более эффективной, если бы ядовита, вероятно, использовалась в качестве защитного оружия маленькими ранними млекопитающими. Она также могла использоваться во время внутривидовая конкуренция или хищничество.[17]

Кости таза Катопсбаатар образец PM120 / 107, возможно, не был сплавлен, потому что слияние произошло на поздней стадии развития, потому что это был сексуально диморфный особенность, встречающаяся только у мужчин (несращенные кости таза могли способствовать расширению родовой канал у женщин), или слияние таза может быть таксономической разницей между Катопсбаатар и другие мультитуберкуляты.[13] В отличие от других млекопитающих, таз мультитуберкулеза очень узкий; у других родов, где известен таз, каждая половина лобка и седалищная кость слились вместе, образуя киль. Длина и жесткость киля указывают на то, что таз не мог расширяться во время родов. Поскольку для прохождения яйца будет мало места (монотремы яйцекладки имеют широкие седалищные дуги), Киелан-Яворовска предположила в 1979 году, что мультитуберкулезные заболевания живородящий (родили живыми), и что новорожденные были очень маленькими, как у сумчатых.[4]
Волосы, структурно похожие на волосы современных млекопитающих и связанные с костями монгольского мультитуберкулеза. Лямбдопсалис, был идентифицирован в копролиты (окаменелые фекалии) хищных млекопитающих из Палеоценовая эпоха. Это указывает на то, что у мультитуберкулятов были волосы для изоляции, как у современных млекопитающих (и, возможно, у ископаемых млекопитающих), что, вероятно, связано с гомеотермия (теплокровность).[4][18]
Кормление и диета
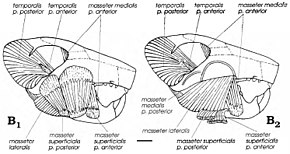
Хотя считалось, что мультитуберкулезные животные были плотоядными или травоядными, с тех пор, как американский палеонтолог Уильям А. Клеменс и Келан-Яворовска предложили современные крысы кенгуру как аналоги для группы в 1979 году они были рассмотрены всеядные (питание как растениями, так и животными).[4] В отличие от млекопитающих у мультитуберкулезных больных применялся обратный жевательный ход, который приводил к жевательные мышцы - мышцы, двигающие нижнюю челюсть, - вставляются больше вперед, чем в других группах (включая грызунов). Гамбарян и Киелан-Яворовска реконструировали жевательную мускулатуру различных мультитуберкулезных больных в 1995 г. и обнаружили, что Катопсбаатар и его родственники обладали очень мощной жевательной мускулатурой из-за их высоких скуловых дуг и больших передних и промежуточных скуловых гребней и венечных отростков. Их мощные резцы с ограниченными полосками эмали были бы хорошо приспособлены для грызения и разрезания твердых семян (подобно грызунам). Поскольку он был больше, чем некоторые другие мультитуберкулезные, Катопсбаатар пришлось бы открывать рот только на 25 градусов, чтобы раздавить твердые семена диаметром 12–14 мм (0,47–0,55 дюйма); разрыв на 40 градусов вызвал бы вывих. После срезания резцов премоляры и моляры начинали шлифоваться «мощным ударом».[12]
По словам Гамбаряна и Келан-Яворовской, приспособление для измельчения твердых семян иногда - например, Катопсбаатар- противодействует преимуществу низкого мыщелкового отростка (что препятствует вывиху нижней челюсти). Передний и промежуточный скуловые гребни черепа были источником поверхностных жевательная мышца, что облегчает жевание. Разделение происхождения этой мышцы на две части и оставленные ими округлые мышечные рубцы уникальны для мультитуберкулезных млекопитающих. Жевательные мышцы мультитуберкулезных независимо развился общие черты с грызунами и мелкими травоядными сумчатыми. Как и у грызунов, мультитуберкуляты могли быть способны к двустороннему жеванию, когда оба ряда зубов нижней челюсти одновременно выполняют одну и ту же функцию, и к одностороннему жеванию (когда используются ряды с одной стороны).[3][12]
Поза и движение

Положение конечностей у мультитуберкулезных больных обсуждалось. По мнению некоторых исследователей, они использовали парасагиттальную стойку с прямыми конечностями под телом; другие считают более вероятной распростертую позу. Киелан-Яворовска и Хурум поддержали последнюю теорию в 2006 году, основываясь на наличии шпор задних ног - особенности, которая, по их мнению, присутствует только у крупных млекопитающих. Они отметили, что все ранние млекопитающие, сохранившиеся в озерный (озерные) отложения были сжаты сверху вниз, что свидетельствует о раскидистой стойке, тогда как более поздние млекопитающие сохранились на их боках. Более ранние аргументы в пользу вытянутой стойки включают глубокий таз и особенности ног. Они также предположили, что ноги мультитуберкулезных стопоходящий (подошва касается земли) в покое, но пальцевидный (подошва не касается земли) при прыжках и быстром беге; они отвергли идею, что передние конечности мультитуберкулезных и других ранних млекопитающих были более парасагиттальными, чем их задние. Келан-Яворовская и Хурум изображены Катопсбаатар со стопоходящими, раскидистыми ногами, с подвижными шпорами, направленными внутрь при подготовке к атаке.[19][4]
В 2008 году Киелан-Яворовска и Хурум предположили, что длинный остистый отросток на Катопсбаатар позвонка и длинные поперечные отростки в Nemegtbaatar может указывать на то, что некоторые мультитуберкулезные больные были скачкообразными (имели способность прыгать). Катопсбаатар вероятно, имел сильные мышцы, прикрепленные к пяточному клубню, что еще раз подтверждает гипотезу о прыжках.[13][20] Хотя предполагалось, что мульти-туберкулезные заболевания древесный (жили на деревьях), большинство азиатских таксонов, вероятно, были наземными; некоторые другие были роющий, копать и жить под землей.[4]
Палеоокружающая среда
Все образцы Катопсбаатар известны из свиты Барун-Гойот пустыни Гоби, которая, вероятно, датируется поздним Кампанский возраст Поздний мел эпоха (около 72 миллионов лет назад). Экземпляры были обнаружены в красных пластах района Эрмиин Цав, за исключением одного коренного зуба из Хулсана. Когда Катопсбаатар Было обнаружено, что красные пласты области Хермиин-Цав считались отдельными формациями, ровесниками (того же геологического возраста) формации Барун-Гойот, поскольку они содержат много одинаковых животных, но теперь считается, что они соответствуют разным уровням свиты Барун-Гойот и Немегт.[3][21][22] Рок фации красных пластов района Эрмиин Цав состоят из оранжевых толстослоистых песчаник, с тонким переплетение светлых иловые камни и аргиллиты.[23][3] Фации горных пород формации Барун-Гойот считаются результатом засушливой или полузасушливой среды с эолийский (отложенные ветром) кровати.[24][25]
Другие известные млекопитающие из красных пластов Хермийн-Цав включают мультитуберкулезных Nemegtbaatar, Чулсанбаатар и Несовбаатар, и терианы Дельтатеридиум, Азиорикты, и Barunlestes.[26] Динозавры включают Ajancingenia, Велоцираптор, Сайхания, Платицератопс, Гобицератопс, и некоторые неопределенные тероподы. Рептилии включают черепаху Монголемис, ящерицы Гобинатус, Чингизавр, Prodenteia, Gladidenagama и Фриносомим, и неопределенный крокодил. Лягушка Gobiates и неопределенный алексорнитиформный птицы также известны. Остракоды включают Лимноцит, Cypridea, и Евкиприс.[27]
Рекомендации
- ^ а б c Kielan-Jaworowska, Z. (1974). «Многотуберкулезная сукцессия в позднем мелу пустыни Гоби (Монголия)» (PDF). Palaeontologica Polonica. Результаты польско-монгольских палеонтологических экспедиций - Часть V. 30: 23–43. В архиве (PDF) из оригинала на 2017-05-05. Получено 2018-03-20.
- ^ Дашзевег, Д .; Novacek, M. J .; Норелл, М. А .; Clark, J.M .; Chiappe, L.M .; Дэвидсон, А .; McKenna, M.C .; Dingus, L .; Swisher, C .; Алтангерел, П. (1995). «Исключительная сохранность нового сообщества позвоночных из позднего мела Монголии». Природа. 374 (6521): 446–449. Bibcode:1995Натура.374..446D. Дои:10.1038 / 374446a0. S2CID 4245878.
- ^ а б c d е ж грамм час я j k л м Kielan-Jaworowska, Z .; Hurum, J. H .; Лопатин, А.В. (2005). "Структура черепа в Катопсбаатар и скуловые гребни у многотуберкулезных млекопитающих ». Acta Palaeontologica Polonica. 50 (5). В архиве из оригинала на 2018-02-24. Получено 2018-03-03.
- ^ а б c d е ж грамм час я j Kielan-Jaworowska, Z .; Cifelli, R.L .; Луо, З.-Х. (2004). Млекопитающие эпохи динозавров: происхождение, эволюция и структура. Нью-Йорк: издательство Колумбийского университета. С. xiv, 249, 260–261, 279, 294–299. ISBN 978-0-231-11918-4.
- ^ Kielan-Jaworowska, Z .; Слоан, Р. Э. (1979). "Катопсалис (Multituberculata) из Азии и Северной Америки и проблема распространения тениолабидид в позднем меловом периоде ». Acta Palaeontologica Polonica. 24 (2): 187–197. В архиве из оригинала на 2018-04-30. Получено 2018-04-29.
- ^ Simmons, N.B .; Десуи, М. (1986). "Парафилия в Катопсалис (Mammalia: Multituberculata) и его биогеографические последствия ". Вклад в геологию, Университет Вайоминга. 24 (3): 87–94. Дои:10.2113 / gsrocky.24.special_paper_3.87.
- ^ а б c Келан-Яворовска, З. (1994). «Новое родовое название для мультитуберкулезного млекопитающего»Джадохтатерий" катопсалоидес". Acta Palaeontologica Polonica. 39 (1): 134–136. В архиве из оригинала на 2018-04-30. Получено 2018-04-29.
- ^ а б Williamson, T. E .; Brusatte, S.L .; Secord, R .; Шелли, С. (2016). «Новый тениолабидоидный мультитуберкулез (Mammalia) из среднего Пуэркана формации Насимиенто, Нью-Мексико, и пересмотр систематики и филогении тениолабидоидов». Зоологический журнал Линнеевского общества. 177 (1): 183–208. Дои:10.1111 / zoj.12336. В архиве из оригинала от 22.09.2017. Получено 2018-10-22.
- ^ Lillegraven, J. A .; Kielan-Jaworowska, Z .; Клеменс, В. А. (1979). Мезозойские млекопитающие: первые две трети истории млекопитающих. Беркли: Калифорнийский университет Press. п. 35. ISBN 978-0520035829.
- ^ Kielan-Jaworowska, Z .; Гамбарян, П. П. (1994). «Посткраниальная анатомия и повадки азиатских многотуберкулезных млекопитающих». Lethaia. 27 (4): 300. Дои:10.1111 / j.1502-3931.1994.tb01578.x.
- ^ а б c Kielan-Jaworowska, Z .; Hurum, J. H .; Карри, П. Дж .; Барсболд Р. (2002). «Новые данные по анатомии многотуберкулезного млекопитающего в позднем меловом периоде». Катопсбаатар". Acta Palaeontologica Polonica. 47 (3): 557–560. В архиве из оригинала на 2018-04-30. Получено 2018-04-29.
- ^ а б c d Гамбарян, П.П .; Келан-Яворовска, З. (1995). «Жевательная мускулатура азиатских тениолабидоидных многотуберкулезных млекопитающих». Acta Palaeontologica Polonica. 40 (1): 45–108. В архиве из оригинала на 2018-02-24. Получено 2018-03-03.
- ^ а б c d е ж грамм Hurum, J. H .; Келан-Яворовска, З. (2008). «Посткраниальный скелет мелового многотуберкулезного млекопитающего. Катопсбаатар". Acta Palaeontologica Polonica. 53 (4): 545–566. Дои:10.4202 / приложение.2008.0401.
- ^ а б c Kielan-Jaworowska, Z .; Хурум, Дж. Х. (1997). «Джадохтатерия - новый подотряд многотуберкулезных млекопитающих». Acta Palaeontologica Polonica. 42 (2): 201–242. В архиве из оригинала на 2018-04-30. Получено 2018-04-29.
- ^ Kielan-Jaworowska, Z .; Хурум, Дж. Х. (2001). «Филогения и систематика многотуберкулезных млекопитающих». Палеонтология. 44 (3): 389–429. Дои:10.1111/1475-4983.00185.
- ^ Kielan-Jaworowska, Z .; Пресли, Р .; Поплин, Сесиль (1986). «Черепная сосудистая система у тениолабидоидных многотуберкулезных млекопитающих». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 313 (1164): 525–602. Bibcode:1986РСПТБ.313..525К. Дои:10.1098 / рстб.1986.0055. JSTOR 2396476.
- ^ а б Hurum, J. H .; Луо, Z-X; Келан-Яворовска, З. (2006). "Были ли млекопитающие изначально ядовитыми?". Acta Palaeontologica Polonica. 51 (1): 1–11. В архиве с оригинала от 27.10.2017. Получено 2018-03-10.
- ^ Meng, J .; Висс, А. Р. (1997). «Волосы мультитуберкулезных и других млекопитающих, извлеченные из экскрементов палеогена». Природа. 385 (6618): 712–714. Bibcode:1997Натура.385..712М. Дои:10.1038 / 385712a0. PMID 9034186. S2CID 4325160.
- ^ Kielan-Jaworowska, Z .; Хурум, Дж. Х. (2006). «Положение конечностей у ранних млекопитающих: раскидистое или парасагиттальное». Acta Palaeontologica Polonica. 51 (3): 393–406. В архиве с оригинала на 2017-10-08. Получено 2018-03-13.
- ^ Chen, M .; Уилсон, Г. П. (2015). «Многофакторный подход к выводу о локомоторных режимах у мезозойских млекопитающих». Палеобиология. 41 (2): 280–312. Дои:10.1017 / pab.2014.14. S2CID 86087687.
- ^ Gradziński, R .; Kielan-Jaworowska, Z .; Марьянская, Т. (1977). «Верхнемеловые отложения Джадохта, Барун-Гойот, верхний мел и Немегт Монголии, включая замечания по предыдущим подразделениям». Acta Geologica Polonica. 7 (3). В архиве из оригинала на 2018-03-14. Получено 2018-03-13.
- ^ Эберт, Д. А. (2018). «Стратиграфия и палеоэкологическая эволюция богатой динозаврами сукцессии Баруунгойот-Немегт (верхний мел), бассейн Немегт, южная Монголия». Палеогеография, палеоклиматология, палеоэкология. 494: 29–50. Bibcode:2018ППП ... 494 ... 29E. Дои:10.1016 / j.palaeo.2017.11.018.
- ^ Gradziński, R .; Ежикевич, Т. (1972). «Дополнительные географические и геологические данные польско-монгольских палеонтологических экспедиций» (PDF). Палеонтология Полоника. 22: 17–32. В архиве (PDF) из оригинала на 2014-08-08. Получено 2018-03-13.
- ^ Eberth, D. A .; Бадамгарав, Д .; Карри, П. Дж. (2009). «Переход Баруунгойот-Немегт (верхний мел) в районе типа Немегт, бассейн Немегт, Южная Центральная Монголия». 고생물 학회지. 25 (1). ISSN 1225-0929. В архиве из оригинала 2018-06-12. Получено 2018-03-10.
- ^ Эберт, Д. А. (2017). «Стратиграфия и палеоэкологическая эволюция богатой динозаврами последовательности Баруунгойот-Немегт (верхний мел), бассейн Немегт, южная Монголия». Палеогеография, палеоклиматология, палеоэкология. 494: 29–50. Bibcode:2018ППП ... 494 ... 29E. Дои:10.1016 / j.palaeo.2017.11.018.
- ^ Kielan-Jaworowska, Z .; Novacek, M. J .; Трофимов, Б. А .; Д., Дашзевег (2000). «Млекопитающие мезозоя Монголии». В Benton, M. J .; Шишкин, М. А .; Анвин, Д. М .; Курочкин, Э. Н. (ред.). Эпоха динозавров в России и Монголии. Кембридж: Издательство Кембриджского университета. С. 573–626. ISBN 978-0521545822.
- ^ Алрой, Дж. «Хермин Цав, Красные пласты [СМПЭ] (ПИН колл. 3142) (мел Монголии)». fossilworks.org. Fossilworks: Gateway to the Paleobiology Database. В архиве из оригинала 14 марта 2018 г.. Получено 13 марта 2018.
внешняя ссылка
 СМИ, связанные с Катопсбаатар в Wikimedia Commons
СМИ, связанные с Катопсбаатар в Wikimedia Commons Данные, относящиеся к Катопсбаатар в Wikispecies
Данные, относящиеся к Катопсбаатар в Wikispecies
| |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||

