Млекопитающее - Mammal


Млекопитающие (из латинский мама "грудь") представляют собой группу позвоночное животное животные составляющий учебный класс Млекопитающие (/мəˈмeɪляə/), и характеризуется наличием молочные железы который в самки производить молоко для кормления (выхаживания) молодняка, неокортекс (область мозга), мех или же волосы, и три кости среднего уха. Эти характеристики отличают их от рептилии и птицы, откуда они расходился в конце Каменноугольный, примерно 300 миллионов лет назад. Около 6400 сохранившийся разновидность млекопитающих были описаны. Самый большой заказы являются грызуны, летучие мыши и Eulipotyphla (ежики, родинки, землеройки, и другие). Следующие три - это Приматы (обезьяны включая люди, обезьяны, и другие), Парнокопытные (китообразные и копытные ), а Хищник (кошки, собаки, уплотнения, и другие).
С точки зрения кладистика, который отражает эволюционную историю, млекопитающие - единственные живые представители Synapsida; это клады, вместе с Sauropsida (рептилии и птицы), составляет большую Амниота клады. Ранние предки синапсидных млекопитающих были сфенакодонт пеликозавры, группа, которая включала немлекопитающих Диметродон. В конце Каменноугольный Около 300 миллионов лет назад эта группа отошла от линии завропсидов, которая привела к сегодняшним рептилиям и птицам. Строка, следующая за стволовая группа Sphenacodontia разделилась на несколько различных групп синапсидов, не относящихся к млекопитающим, иногда ошибочно называемых рептилиями, подобными млекопитающим, прежде чем дать начало Терапсида в Ранняя пермь период. Современные отряды млекопитающих возникли в Палеоген и Неоген периоды Кайнозойский эра, после вымирание нептичьих динозавров, и были доминирующей группой наземных животных с 66 миллионов лет назад по настоящее время.
Основной тип телосложения - четвероногий, и большинство млекопитающих используют свои четыре конечности для наземное движение; но у некоторых конечности приспособлены для жизнь на море, в воздух, на деревьях, под землей, или же на двух ногах. Размер млекопитающих варьируется от 30 до 40 мм (1,2–1,6 дюйма). летучая мышь шмеля до 30 м (98 футов) синий кит - возможно, самое большое животное из когда-либо существовавших. Максимальный срок службы варьируется от двух лет для землеройка до 211 лет для гренландский кит. Все современные млекопитающие рождают живых детенышей, кроме пяти разновидность из монотремы, которые являются откладывающими яйца млекопитающими. Самая богатая видами группа млекопитающих, когорта называется плаценты, есть плацента, что позволяет кормить плод во время беременность.
Большинство млекопитающих разумный, у некоторых из них большой мозг, самосознание, и использование инструмента. Млекопитающие могут общаться и вокализировать несколькими способами, включая УЗИ, ароматическая маркировка, сигналы тревоги, пение, и эхолокация. Млекопитающие могут организовываться в общества ядерного синтеза, гаремы, и иерархии - но также может быть одиночным и территориальный. Большинство млекопитающих полигинный, но некоторые могут быть моногамный или же полиандрический.
Одомашнивание многих видов млекопитающих люди играли важную роль в Неолитическая революция, и в результате сельское хозяйство замена охота и собирательство как основной источник пищи для человека. Это привело к серьезной перестройке человеческих обществ от кочевых к оседлым, с более тесным сотрудничеством между все более и более многочисленными группами и, в конечном итоге, к развитию первых цивилизации. Домашние млекопитающие обеспечивали и продолжают обеспечивать энергией транспорт и сельское хозяйство, а также пищу (мясо и молочные продукты ), мех, и натуральная кожа. Млекопитающие также охотился и гонялись для спорта, и используются как модельные организмы в науке. Млекопитающие изображены в Изобразительное искусство поскольку Палеолит раз, и появляются в литературе, кино, мифологии и религии. Снижение численности и вымирание многих млекопитающих главным образом управляется человеком браконьерство и разрушение среды обитания, в первую очередь вырубка леса.
Классификация

Классификация млекопитающих прошла несколько итераций с тех пор, как Карл Линней изначально определил класс. Общепринятой системы классификации не существует; Маккенна и Белл (1997) и Уилсон и Ридер (2005) предоставляют полезные недавние сборники.[1] Джордж Гейлорд Симпсон "Принципы классификации и классификации млекопитающих" (AMNH Бюллетень v. 85, 1945) предусматривает систематика о происхождении и взаимоотношениях млекопитающих, которым повсеместно учили до конца 20 века. Начиная с классификации Симпсона, палеонтологическая запись был перекалиброван, и за прошедшие годы было много споров и прогресса в отношении теоретических основ систематизации как таковой, отчасти благодаря новой концепции кладистика. Хотя полевые исследования постепенно сделали классификацию Симпсона устаревшей, она остается наиболее близкой к официальной классификации млекопитающих.[2]
Большинство млекопитающих, включая шесть самых богатых видами заказы, принадлежат к плацентарной группе. Три крупнейших отряда по количеству видов: Rodentia: мышей, крысы, дикобразы, бобры, капибары и другие грызущие млекопитающие; Рукокрылые: летучие мыши; и Сорикоморфа: землеройки, родинки и соленодоны. Следующие три крупнейших заказа, в зависимости от биологическая классификация схема используется, являются Приматы в том числе обезьяны, обезьяны и лемуры; то Cetartiodactyla включая киты и копытные; и Хищник который включает кошки, собаки, ласки, медведи, уплотнения и союзники.[3] В соответствии с Виды млекопитающих мира В 2006 г. было идентифицировано 5416 видов. Они были сгруппированы в 1229 видов. роды, 153 семьи и 29 орденов.[3] В 2008 г. Международный союз охраны природы (МСОП) завершил пятилетнюю Глобальную оценку млекопитающих Красный список МСОП, в котором насчитывалось 5 488 видов.[4] Согласно исследованию, опубликованному в Журнал маммологии В 2018 году число признанных видов млекопитающих составило 6495, включая 96 недавно исчезнувших.[5]
Определения
Слово "млекопитающее "современно, от научного названия Млекопитающие изобретен Карлом Линнеем в 1758 году, происходит от латинский мама ("сосок, папа"). Во влиятельной статье 1988 года Тимоти Роу дал определение Mammalia филогенетически как группа короны млекопитающих, клады состоящий из самый последний общий предок жизни монотремы (ехидны и утконос ) и Териан млекопитающие (сумчатые и плаценты ) и всех потомков этого предка.[6] Поскольку этот предок жил в Юрский период, определение Роу исключает всех животных из более раннего Триасовый, несмотря на то, что окаменелости триаса в Харамийида относятся к млекопитающим с середины 19 века.[7] Если рассматривать Mammalia как коронную группу, его происхождение можно приблизительно датировать как первое известное появление животных, более тесно связанных с одними существующими млекопитающими, чем с другими. Амбондро более тесно связан с монотремами, чем с терианскими млекопитающими, в то время как Амфилетес и Амфитерий более тесно связаны с терианами; окаменелости всех трех родов датируются примерно 167 миллион лет назад в Средняя юра, это разумная оценка появления коронной группы.[8]
Т. С. Кемп дал более традиционное определение: "Синапсиды которые обладают зубной –чешуйчатый сочленение челюсти и окклюзия между верхними и нижними молярами с поперечным компонентом движения »или, что эквивалентно, по мнению Кемпа, клады, происходящие от последнего общего предка Sinoconodon и живые млекопитающие.[9] Самый ранний известный синапсид, удовлетворяющий определениям Кемпа, - это Тикитериум, датированный 225 Ма, поэтому внешний вид млекопитающих в этом более широком смысле можно описать как Поздний триас Дата.[10][11]
Классификация Маккенны / Белла
В 1997 г. млекопитающие были полностью пересмотрены Малкольм С. МакКенна и Сьюзен К. Белл, что привело к классификации Маккенны / Белла. Их книга 1997 года, Классификация млекопитающих выше видового уровня,[12] представляет собой всеобъемлющую работу по систематике, взаимосвязям и встречаемости всех таксонов млекопитающих, живых и вымерших, вплоть до ранга родов, хотя молекулярно-генетические данные бросают вызов некоторым группировкам более высокого уровня. Авторы работали вместе как палеонтологи на Американский музей естественной истории, Нью-Йорк. Маккенна унаследовал проект от Симпсона и вместе с Беллом построил полностью обновленную иерархическую систему, охватывающую живые и вымершие таксоны, которая отражает историческую генеалогию млекопитающих.[2]
В следующем списке вымерший группы помечены значком кинжал (†).
Класс Mammalia
- Подкласс Прототерия: monotremes: ехидны и утконос
- Подкласс Тереобразные: живородящие млекопитающие и их доисторические сородичи.
- Инфракласс †Аллотерия: мультитуберкулезные
- Инфракласс †Eutriconodonta: eutriconodonts
- Инфракласс Голотерия: современные живородящие млекопитающие и их доисторические сородичи.
- Суперлегион †Куехнеотерия
- Суперкогорта Терия: живородящие млекопитающие
- Когорта Marsupialia: сумчатые
- Магнордер Австралиделфия: Австралийские сумчатые и Монито дель Монте
- Магнордер Америдельфия: Сумчатые животные Нового Света. Теперь считается парафилетическим, с опоссумы землеройки быть ближе к австралийцам.[13]
- Когорта Плаценталия: плаценты
- Магнордер Ксенартра: xenarthrans
- Магнордер Эпитерия: epitheres
- Суперзаказ †Лептиктида
- Суперзаказ Прептотерия
- Grandorder Анагалида: зайцеобразные, грызуны и слоновьи землеройки
- Grandorder Ferae: плотоядные, панголины, †креодонты и родственники
- Grandorder Липотифла: насекомоядные
- Grandorder Archonta: летучие мыши, приматы, Colugos и деревья
- Grandorder Унгулата: копытные
- Заказ Tubulidentata incertae sedis: трубкозуб
- Зеркало Eparctocyona: †Condylarths, киты и парнокопытные (копытные животные)
- Зеркало †Меридиунгулата: Южноамериканские копытные.
- Зеркало Altungulata: периссодактилии (однопалые копытные), слоны, ламантины и даманы
- Когорта Marsupialia: сумчатые
Молекулярная классификация плаценты
В начале 21 века молекулярные исследования, основанные на ДНК анализ предложил новые отношения между семьями млекопитающих. Большинство этих результатов были независимо подтверждены ретротранспозон данные о присутствии / отсутствии.[14] Системы классификации, основанные на молекулярных исследованиях, выявляют три основные группы или линии плацентарных млекопитающих:Афротерия, Ксенартра и Бореевтерия -который расходился в Меловой. Отношения между этими тремя линиями передачи спорны, и были предложены все три возможные гипотезы относительно того, какая группа базальный. Эти гипотезы Атлантогената (базальная бореэвтерия), Эпитерия (базальная ксенартра) и Экзафроплаценталия (базальная афротерия).[15] Boreoeutheria, в свою очередь, состоит из двух основных линий:Euarchontoglires и Лавразиатерия.
Оценки времени расхождения между этими тремя группами плаценты колеблются от 105 до 120 миллионов лет назад, в зависимости от типа используемой ДНК (например, ядерный или же митохондриальный )[16] и различные интерпретации палеогеографический данные.[15]
| Млекопитающие |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

В кладограмма выше основано на Тарвере и другие. (2016)[17]
Группа I: Суперзаказ Афротерия[18]
- Clade Афроинсектифилия
- Заказ Macroscelidea: слоновьи землеройки (Африка)
- Заказ Афросорицида: тенреки и золотые родинки (Африка)
- Заказ Tubulidentata: трубкозуб (Африка к югу от Сахары)
- Clade Паенунгулата
- Заказ Hyracoidea: даманы или дассии (Африка, Аравия)
- Заказ Хоботок: слоны (Африка, Юго-Восточная Азия)
- Заказ Сирения: дюгони и ламантины (космополитичный тропический)
Группа II: Суперзаказ Ксенартра[18]
- Заказ Pilosa: ленивцы и муравьеды (неотропики)
- Заказ Cingulata: броненосцы и вымершие родственники (Америка)
Группа III: Magnaorder Бореевтерия[18]
- Суперзаказ: Euarchontoglires (Супраприматы )
- Grandorder Euarchonta
- Заказ Scandentia: treehrews (Юго-Восточная Азия).
- Заказ Dermoptera: летающие лемуры или колуго (Юго-Восточная Азия)
- Заказ Приматы: лемуры, кустарники, обезьяны, обезьяны, люди (космополитичный)
- Grandorder Glires
- Заказ Зайцеобразные: пищухи, кролики, зайцы (Евразия, Африка, Америка)
- Заказ Rodentia: грызуны (космополиты)
- Grandorder Euarchonta
- Суперзаказ: Лавразиатерия
- Заказ Eulipotyphla: землеройки, ежи, кроты, соленодоны
- Clade Скротифера
- Заказ Рукокрылые: летучие мыши (космополитичный)
- Clade Fereuungulata
- Clade Ferae
- Clade Euungulata
- Заказ Cetartiodactyla: китообразные (киты, дельфины и морские свиньи) и копытные, в том числе свиньи, крупный рогатый скот, олень и жирафы
- Заказ Периссодактиля: четвероногие копытные, в том числе лошади, ослы, зебры, тапиры и носороги
Эволюция
Происхождение
Synapsida, клады, в которой содержатся млекопитающие и их вымершие родственники, возникла во время Пенсильванский подпериод (От ~ 323 миллионов до ~ 300 миллионов лет назад), когда они отделились от рептилий и птиц. Млекопитающие группы кроны произошли от более ранних млекопитающие вовремя Раннеюрский период. Кладограмма показывает, что Mammalia является коронной группой.[19]
| Млекопитающие |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Эволюция от амниот

Первый полностью земной позвоночные мы амниот. Как их амфибия четвероногий у предшественников были легкие и конечности. Однако у амниотических яиц есть внутренние оболочки, которые позволяют развиваться эмбрион дышать, но сохранять воду. Следовательно, амниоты могут откладывать яйца на суше, в то время как амфибии обычно откладывают яйца в воде.
Первые амниоты, по-видимому, возникли в пенсильванский субпериод Каменноугольный. Они произошли от ранее рептилиоморф амфибии четвероногие,[20] которые жили на земле, которая уже была заселена насекомые и других беспозвоночных, а также папоротники, мхи и другие растения. В течение нескольких миллионов лет были выделены две важные линии передачи амниот: синапсиды, который позже будет включать общего предка млекопитающих; и завропсиды, которые теперь включают черепахи, ящерицы, змеи, крокодилы и динозавры (включая птицы ).[21] Синапсиды имеют одно отверстие (височное окно ) низко с каждой стороны черепа. Одна группа синапсидов, пеликозавры, включал в себя самых крупных и свирепых животных раннего Пермский период.[22] Синапсиды, не относящиеся к млекопитающим, иногда (неточно) называют «рептилиями, подобными млекопитающим».[23][24]
Терапсиды, группа синапсидов, произошедшая от пеликозавры в Средняя пермь, около 265 миллионов лет назад, и стали доминирующими наземными позвоночными.[23] Они отличаются от базальных евпеликозавры в нескольких особенностях черепа и челюстей, в том числе: более крупные черепа и резцы равных по размеру у терапсид, но не у европеликозавров.[23] Линия терапсид, ведущая к млекопитающим, прошла ряд стадий, начиная с животных, очень похожих на своих предков пеликозавров, и заканчивая пробаиногнат цинодонты, некоторые из которых можно легко принять за млекопитающих. Эти этапы характеризовались:[25]
- Постепенное развитие костной вторичной вкус.
- Переход к вертикальной позе конечностей, что повысит выносливость животных, избегая Ограничение перевозчика. Но этот процесс был медленным и беспорядочным: например, у всех травоядных терапевсидов, не относящихся к млекопитающим, сохранились раскидистые конечности (у некоторых поздних форм могли быть полуверстые задние конечности); Пермские хищные терапсиды имели распростертые передние конечности, а у некоторых позднепермских также были полурасположенные задние конечности. Фактически, у современных монотремов все еще есть полусвободные конечности.
- В зубной постепенно стала основной костью нижней челюсти, которая к триасу переросла в челюсть полностью млекопитающих (нижняя состоит только из зубной кости) и среднее ухо (которое состоит из костей, которые ранее использовались для построения челюстей рептилий). ).
Первые млекопитающие
В Пермско-триасовое вымирание около 252 миллионов лет назад, что было продолжительным событием из-за накопления нескольких импульсов вымирания, положило конец господству плотоядных терапсид.[26] В раннем триасе большинство ниш средних и крупных наземных хищников заняли архозавры[27] которые за длительный период (35 миллионов лет) стали включать крокодиломорфы,[28] то птерозавры и динозавры;[29] однако крупные цинодонты, такие как Труцидоцинодон и траверсодонтиды по-прежнему занимали ниши крупных хищников и травоядных соответственно. К юрскому периоду динозавры также стали доминировать в нишах крупных наземных травоядных.[30]
Первые млекопитающие (в понимании Кемпа) появились в эпоху позднего триаса (около 225 миллионов лет назад), через 40 миллионов лет после появления первых терапсид. Они вышли из своей ночной жизни насекомоядное животное ниша с середины юры и далее;[31] Юрский период Castorocauda, например, был близким родственником настоящих млекопитающих, имевших приспособления для плавания, рытья и ловли рыбы.[32] Считается, что большинство, если не все, вели ночной образ жизни ( ночное узкое место ), что составляет большую часть типичных черт млекопитающих.[33] Большинство видов млекопитающих, существовавших в Мезозойская эра были мультитуберкулезными, эутриконодонтами и спалакотерииды.[34] Самый ранний из известных метатериан является Sinodelphys, найденный в 125-миллионном возрасте Раннемеловой период сланец на северо-востоке Китая Провинция Ляонин. Ископаемое почти готово и включает пучки меха и отпечатки мягких тканей.[35]

Самая старая известная окаменелость среди Eutheria («настоящих зверей») - это маленькие землеройки. Juramaia sinensis, или «юрская мать из Китая», датируемая 160 миллионами лет назад в конце юрского периода.[36] Более поздний родственник-евтериец, Eomaia, датируемый 125 миллионами лет назад в раннем меловом периоде, обладал некоторыми общими чертами с сумчатыми, но не с плацентарными, что свидетельствует о том, что эти особенности присутствовали у последнего общего предка этих двух групп, но позже были потеряны в плацентарной линии.[37] В частности, надлобковые кости простираться вперед от таза. Они не встречаются ни в каких современных плацентарных животных, но встречаются у сумчатых, одноплодных, других млекопитающих, не относящихся к териям, и Ухаатерий, раннее меловое животное в эвтерианском отряде Азиориктитерия. Это касается и мультитуберкулятов.[38] Они, по-видимому, являются наследственным признаком, который впоследствии исчез в плацентарной линии. Эти надлобковые кости, по-видимому, функционируют, укрепляя мышцы во время передвижения, уменьшая количество представленного пространства, которое требуется плацентарям для удержания их плод в период беременности. Узкий выход из таза указывает на то, что детеныши были очень маленькими при рождении и поэтому беременность был коротким, как у современных сумчатых. Это говорит о том, что плацента была более поздним развитием.[39]
Одна из самых ранних известных монотрем была Тейнолофос, который жил около 120 миллионов лет назад в Австралии.[40] У монотрем есть некоторые особенности, которые могут быть унаследованы от исходных амниот, например, одно и то же отверстие для мочеиспускания, дефекации и размножения (клоака ) —Как делают ящерицы и птицы—[41] и они лежат яйца кожистые и не кальцинированные.[42]
Самые ранние появления функций
Адрокодиум, окаменелости которых датируются примерно 195 миллионов лет назад, в раннем Юрский, представляет собой первое четкое свидетельство челюстного сустава, образованного исключительно чешуйчатой и зубной костями; в челюсти нет места для сустава, кости, входящей в состав челюстей всех ранних синапсидов.[43]

Самые ранние явные свидетельства наличия волос или меха находятся в окаменелостях Castorocauda и Мегаконус, от 164 миллионов лет назад в середине юрского периода. В 1950-х годах было высказано предположение, что отверстия (проходы) в максиллы и предчелюстные кости (кости в передней части верхней челюсти) цинодонтов были каналами, которые снабжали кровеносные сосуды и нервы к вибриссам (усы ) и также были доказательства волос или меха;[44][45] однако вскоре было отмечено, что отверстия не обязательно указывают на наличие вибрисс у животного, как современная ящерица. Тупинамбис имеет отверстия, которые почти идентичны тем, которые встречаются у цинодонта, не являющегося млекопитающим Тринаксодон.[24][46] Популярные источники, тем не менее, продолжают относить усы к Тринаксодон.[47] Исследования перми копролиты предполагают, что не млекопитающие синапсиды эпохи уже был мех, что указывает на эволюцию волос, возможно, еще дицинодонты.[48]
Когда эндотермия впервые появился в эволюции млекопитающих, остается неясным, хотя, как правило, считается, что он впервые появился у не млекопитающих. терапсиды.[48][49] Современные монотремы имеют более низкую температуру тела и более изменчивую скорость метаболизма, чем сумчатые и плацентарные.[50] но есть свидетельства того, что некоторые из их предков, включая, возможно, предков териан, могли иметь температуру тела, как у современных териан.[51] Точно так же у некоторых современных териан, таких как афротеры и ксенартранцы, вторично снизилась температура тела.[52]
Эволюция стоячих конечностей у млекопитающих не завершена - у живых и ископаемых монотрем конечности распростерлись. Парасагиттальная (не разгибающаяся) поза конечностей появилась где-то в конце юрского или раннемелового периода; это найдено в евтерианском Eomaia и метатериан Sinodelphys, оба датированы 125 миллионами лет назад.[53] Надлобковый кости, особенность, которая сильно повлияла на размножение большинства клад млекопитающих, впервые были обнаружены в Tritylodontidae, предполагая, что это синапоморфия между ними и млекопитающее. Они вездесущи у неплацентарных млекопитающих, хотя Мегазостродон и Эритротерий похоже, их не хватало.[54]
Было высказано предположение, что первоначальная функция кормление грудью (молоко продукция) заключалась в том, чтобы яйца оставались влажными. Большая часть аргументов основана на монотремах, млекопитающих, откладывающих яйца.[55][56]
Восстание млекопитающих
Терианские млекопитающие заняли средние и крупные экологические ниши в Кайнозойский, после Меловое – палеогеновое вымирание приблизительно 66 миллионов лет назад опустело экологическое пространство, когда-то заполненное нептичьими динозаврами и другими группами рептилий, а также различными другими группами млекопитающих,[57] и претерпел экспоненциальное увеличение размеров тела (мегафауна ).[58] Затем млекопитающие очень быстро изменились; как птицы, так и млекопитающие демонстрируют экспоненциальный рост разнообразия.[57] Например, самая ранняя из известных летучих мышей датируется примерно 50 миллионами лет назад, всего через 16 миллионов лет после исчезновения нептичьих динозавров.[59]
Молекулярно-филогенетические исследования первоначально предполагали, что большинство порядков плаценты разошлись примерно от 100 до 85 миллионов лет назад и что современные семьи появились в период с позднего времени. эоцен сквозь Миоцен.[60] Однако окаменелости плаценты до конца мелового периода не обнаружены.[61] Самые ранние бесспорные окаменелости плаценты относятся к раннему Палеоцен, после исчезновения нептичьих динозавров.[61] В частности, ученые определили раннее палеоценовое животное по имени Protungulatum donnae как одно из первых плацентарных млекопитающих.[62] однако он был реклассифицирован как бесплацентарный эуттерианский.[63] Повторная калибровка показателей генетического и морфологического разнообразия позволила предположить Поздний мел происхождение плацентарных и палеоценовое происхождение большинства современных клад.[64]
Самый ранний из известных предков приматов - Archicebus ахиллес[65] примерно 55 миллионов лет назад.[65] Этот крошечный примат весил 20–30 граммов (0,7–1,1 унции) и мог поместиться на ладони человека.[65]
Анатомия
Отличительные черты
Живые виды млекопитающих можно идентифицировать по наличию потовые железы, включая те, которые специализируются на производстве молока для питания своих молодых.[66] Однако при классификации окаменелостей необходимо использовать другие признаки, поскольку железы мягких тканей и многие другие особенности не видны на окаменелостях.[67]
Многие черты, присущие всем живым млекопитающим, проявились у самых ранних членов группы:
- Челюстной сустав - The зубной (кость нижней челюсти, несущая зубы) и чешуйчатый (маленький черепной кость) встречаются, образуя сустав. В большинстве гнатомы, в том числе ранние терапсиды сустав состоит из суставной (небольшая кость в задней части нижней челюсти) и квадратный (небольшая кость в задней части верхней челюсти).[43]
- Среднее ухо - У млекопитающих кроновой группы звук передается от барабанная перепонка цепочкой из трех костей молоточек, то наковальня и стремени. Изначально молоток и наковальня произошли от суставной и квадратной костей, которые составляли челюстной сустав ранних терапсид.[68]
- Замена зуба - Зубы можно заменить один раз (дифиодонтия ) или (как у зубатых китов и мюрид грызуны) совсем нет (монофиодонтия ).[69] У слонов, ламантинов и кенгуру на протяжении всей жизни постоянно появляются новые зубы (полифиодонтия ).[70]
- Призматическая эмаль - The эмаль покрытие на поверхности зуба состоит из призм, твердых стержневидных структур, отходящих от дентин к поверхности зуба.[71]
- Затылочные мыщелки - Два выступа у основания черепа подходят к самому верхнему шейный позвонок; большинство других четвероногие, напротив, имеют только одну такую ручку.[72]
По большей части эти характеристики отсутствовали у триасовых предков млекопитающих.[73] Почти все млекопитающие имеют надлобковую кость, за исключением современной плаценты.[74]
Половой диморфизм
В среднем самцы млекопитающих крупнее самок, причем самцы как минимум на 10% крупнее самок более чем у 45% исследованных видов. Большинство отрядов млекопитающих также демонстрируют половой диморфизм, ориентированный на самцов, хотя некоторые отряды не обнаруживают какой-либо предвзятости или сильно ориентированы на самок (Lagomorpha). Половой диморфизм размера увеличивается с размером тела у млекопитающих (правило Ренша), что позволяет предположить, что существует параллельное давление отбора как на мужской, так и на женский размер. Диморфизм, ориентированный на самцов, относится к половому отбору самцов через конкуренцию самцов за самок, так как существует положительная корреляция между степенью полового отбора, на что указывают системы спаривания, и степенью диморфизма размеров, ориентированного на самцов. Степень полового отбора также положительно коррелирует с размерами самцов и самок у млекопитающих. Кроме того, параллельное давление отбора на массу самок идентифицируется в том возрасте, когда отлучение от груди значительно выше у более полигинных видов, даже с поправкой на массу тела. Кроме того, скорость размножения ниже у более крупных самок, что указывает на то, что отбор по плодовитости у млекопитающих выбирает более мелких самок. Хотя эти закономерности характерны для млекопитающих в целом, существуют значительные различия между отрядами.[75]
Биологические системы
У большинства млекопитающих семь шейные позвонки (кости шеи), в том числе летучие мыши, жирафы, киты и люди. Исключение составляют ламантин и двупалый ленивец, которых всего шесть, а трехпалый ленивец который имеет девять шейных позвонков.[76] Мозг всех млекопитающих обладает неокортекс, область мозга, уникальная для млекопитающих.[77] У плацентарных млекопитающих мозолистое тело, в отличие от одноцветных и сумчатых.[78]
В легкие у млекопитающих губчатые и сотовые. Дыхание в основном достигается с помощью диафрагма, который отделяет грудную клетку от брюшной полости, образуя выпуклый к грудной клетке купол. Сокращение диафрагмы сглаживает купол, увеличивая объем легочной полости. Воздух поступает через ротовую и носовую полости и проходит через гортань, трахею и бронхи, и расширяет альвеолы. Расслабление диафрагмы имеет противоположный эффект, уменьшая объем легочной полости, в результате чего воздух выталкивается из легких. Во время тренировки брюшная стенка контракты, увеличивая давление на диафрагму, в результате чего воздух выходит быстрее и сильнее. В грудная клетка способен расширять и сокращать грудную полость под действием других дыхательных мышц. Следовательно, воздух всасывается или выталкивается из легких, всегда опускаясь по градиенту давления.[79][80] Этот тип легкого известен как легкое мехов из-за его сходства с кузнечным. мехи.[80]
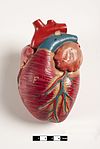
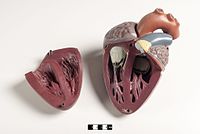
Млекопитающее сердце имеет четыре камеры, две верхние предсердие, приемные камеры и две нижние желудочки, разгрузочные камеры.[81] Сердце имеет четыре клапана, которые разделяют его камеры и обеспечивают кровоток в правильном направлении через сердце (предотвращая обратный ток). После газообмен в легочных капиллярах (кровеносных сосудах в легких) богатая кислородом кровь возвращается в левое предсердие через один из четырех легочные вены. Кровь почти непрерывно течет обратно в предсердие, которое действует как приемная камера, а отсюда через отверстие в левый желудочек. Большая часть крови проходит в сердце пассивно, в то время как предсердия и желудочки расслаблены, но ближе к концу период релаксации желудочков левое предсердие сократится, накачивая кровь в желудочек. Сердце также требует питательных веществ и кислорода, содержащихся в крови, как и другие мышцы, и снабжается им через коронарные артерии.[82]


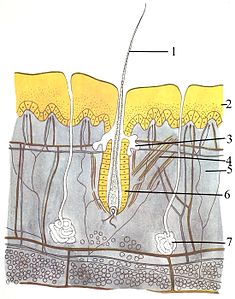
В покровная система (кожа) состоит из трех слоев: самый верхний эпидермис, то дерма и гиподерма. Толщина эпидермиса обычно составляет от 10 до 30 клеток; его основная функция - обеспечение водонепроницаемого слоя. Его внешние клетки постоянно теряются; его самые нижние клетки постоянно делятся и продвигаются вверх. Средний слой, дерма, в 15-40 раз толще эпидермиса. Дерма состоит из многих компонентов, таких как костные структуры и кровеносные сосуды. Гиподерма состоит из жировая ткань, который накапливает липиды и обеспечивает амортизацию и изоляцию. Толщина этого слоя широко варьируется от вида к виду;[83]:97 морские млекопитающие требуется толстая подкожная клетчатка (ворвань ) для изоляции, и правые киты самый толстый жир - 20 дюймов (51 см).[84] Хотя у других животных есть такие особенности, как усы, перья, щетинки, или же реснички внешне напоминающие его, у животных, кроме млекопитающих, нет волосы. Это окончательная характеристика класса. Хотя у некоторых млекопитающих их очень мало, тщательное изучение выявляет характерные черты, часто в неясных частях их тел.[83]:61


Травоядные животные развили широкий спектр физических структур для облегчения потребление растительного материала. Чтобы разрушить неповрежденные ткани растений, млекопитающие разработали зубы структуры, отражающие их пищевые предпочтения. Например, плодоядные (животные, которые питаются в основном фруктами) и травоядные, которые питаются мягкой листвой, имеют зубы с низкой коронкой, специально предназначенные для измельчения листвы и семена. Выпас животные, которые любят много есть, кремнезем - богатые травы, имеют зубы с высокой коронкой, которые способны измельчать твердые ткани растений и не изнашиваются так быстро, как зубы с низкой коронкой.[85] Большинство плотоядных млекопитающих имеют Carnassialiforme зубы (разной длины в зависимости от диеты), длинные клыки и аналогичные схемы замены зубов.[86]
Желудок Парнокопытные разделен на четыре части: рубец, то сеточка, то омазум и сычуг (Только жвачные животные есть рубец). После употребления растительного материала он смешивается со слюной в рубце и сетке и разделяется на твердый и жидкий материал. Твердые частицы слипаются, образуя болюс (или же жвачка ), и его срыгивают. Когда болюс попадает в рот, жидкость выдавливается языком и снова проглатывается. Проглоченная пища попадает в рубец и сетку, где клетчатка микробы (бактерии, простейшие и грибы ) производить целлюлаза, что необходимо для разбивки целлюлоза в растениях.[87] Периссодактили, в отличие от жвачных, хранят переваренную пищу, вышедшую из желудка, в увеличенном виде. слепая кишка, где он ферментируется бактериями.[88] У плотоядных животных простой желудок, приспособленный для переваривания в первую очередь мяса, в отличие от сложных пищеварительных систем травоядных животных, которые необходимы для расщепления жестких и сложных растительных волокон. В слепая кишка либо отсутствует, либо короткий и простой, а толстый кишечник не саккулированный или намного шире тонкой кишки.[89]

Млекопитающее выделительная система включает в себя множество компонентов. Как и большинство других наземных животных, млекопитающие уреотелический, и конвертировать аммиак в мочевина, что делается печень как часть цикл мочевины.[90] Билирубин, отходы производства кровяные клетки, проходит через желчь и моча с помощью ферментов выводится печенью.[91] Прохождение билирубина с желчью через кишечного тракта дает млекопитающим кал характерная коричневая окраска.[92] Отличительные особенности почек млекопитающих включают наличие почечная лоханка и почечные пирамиды, и четко различимого кора и мозговое вещество, что связано с наличием удлиненной петли Генле. Бобовидную форму имеют только почки млекопитающих, хотя есть и исключения, например, многодольчатая. почки почки ластоногих, китообразные и медведи.[93][94] У большинства взрослых плацентарных млекопитающих не остается следов клоака. В эмбрионе эмбриональная клоака делится на заднюю область, которая становится частью ануса, и переднюю область, которая имеет разные судьбы в зависимости от пола человека: у женщин она развивается в преддверие, которое получает уретра и влагалище, а у самцов он образует всю половой член уретра.[94] Тем не менее тенреки, золотые родинки, и немного землеройки сохраняют клоаку как взрослые.[95] У сумчатых половые пути отделены от анального отверстия, но след исходной клоаки остается снаружи.[94] Monotremes, что переводится с Греческий в «единственное отверстие», есть настоящая клоака.[96]
Звуковое производство

Как и у всех других четвероногих, у млекопитающих есть гортань который может быстро открываться и закрываться, чтобы издавать звуки, а надглазничный голосовой тракт который фильтрует этот звук. Легкие и окружающая их мускулатура обеспечивают поток воздуха и давление, необходимые для фонат. Гортань контролирует подача и объем звука, но сила легких выдох также способствует увеличению объема. Более примитивные млекопитающие, такие как ехидна, могут только шипеть, поскольку звук достигается исключительно путем выдоха через частично закрытую гортань. Фонаты других млекопитающих голосовые складки, в отличие от голосовых связок, наблюдаемых у птиц и рептилий. Движение или напряжение голосовых связок может приводить к появлению множества звуков, например мурлыканье и кричать. Млекопитающие могут изменять положение гортани, позволяя им дышать через нос, глотая через рот, и образовывать как оральный, так и носовой звуки; носовые звуки, такие как вой собаки, обычно являются тихими звуками, а звуки рта, например лай собаки, обычно громкими.[97]
У некоторых млекопитающих большая гортань и, следовательно, низкий голос, а именно летучая мышь с головой молотка (Hypsignathus monstrosus), где гортань может занимать всю грудная полость толкая легкие, сердце и трахею в брюшная полость.[98] Большие голосовые пэды также могут снизить высоту звука, как в низком реве большие кошки.[99] Производство инфразвук возможно у некоторых млекопитающих, таких как Африканский слон (Локсодонта spp.) и усатые киты.[100][101] Мелкие млекопитающие с маленькой гортани способны производить УЗИ, который можно обнаружить, изменив среднее ухо и улитка. Ультразвук не слышен для птиц и рептилий, что могло иметь важное значение в мезозой, когда птицы и рептилии были доминирующими хищниками. Этот частный канал используется некоторыми грызунами, например, при общении матери с щенком, а также летучими мышами при эхолокации. Зубастые киты также используют эхолокацию, но, в отличие от голосовой перепонки, которая идет вверх от голосовых связок, у них есть дыня управлять звуками. У некоторых млекопитающих, а именно у приматов, к гортани прикреплены воздушные мешки, которые могут снижать резонансы или увеличивать громкость звука.[97]
Система вокального производства контролируется ядра черепных нервов в мозгу, и снабжается возвратный гортанный нерв и верхний гортанный нерв, филиалы блуждающий нерв. Голосовой тракт снабжен подъязычный нерв и лицевые нервы. Электрическая стимуляция периакведуктальный серый (PEG) область млекопитающих средний мозг вызывать вокализацию. Способность изучать новые вокализации проявляется только у людей, тюленей, китообразных, слонов и, возможно, летучих мышей; у людей это результат прямой связи между моторная кора, который управляет движением, а двигательные нейроны в спинном мозге.[97]
Мех

Основная функция меха млекопитающих - терморегуляция. Другие включают защиту, сенсорные цели, гидроизоляцию и маскировку.[102] Разные виды меха служат разным целям:[83]:99
- Окончательный - что может быть пролить после достижения определенной длины
- Вибриссы - чувствительные волоски, чаще всего усы
- Пелаге - остевой волос, подшерсток и волосы ости
- Колючки - жесткий остевой волос, используемый для защиты (например, в дикобразы )
- Щетина - длинные волосы, обычно используемые в визуальных сигналах. (например, львиный грива )
- Велли - часто называемый «пуховый мех», изолирующий новорожденных млекопитающих.
- Шерсть - длинные, мягкие и часто кудрявые
Терморегуляция
Длина волос не является фактором терморегуляции: например, у некоторых тропических млекопитающих, таких как ленивцы, длина шерсти такая же, как у некоторых арктических млекопитающих, но с меньшей изоляцией; и, наоборот, другие тропические млекопитающие с короткой шерстью обладают такой же изоляционной способностью, как арктические млекопитающие. Плотность меха может повысить теплоизоляционные свойства животных, особенно у арктических млекопитающих густой мех; например, овцебык Имеет остевой волос размером 30 см (12 дюймов), а также плотный подшерсток, который образует воздухонепроницаемую шерсть, позволяющую им выживать при температурах -40 ° C (-40 ° F).[83]:162–163 Некоторые пустынные млекопитающие, такие как верблюды, используют густой мех, чтобы солнечное тепло не достигало их кожи, позволяя животным сохранять прохладу; мех верблюда может достигать 70 ° C (158 ° F) летом, но кожа остается при 40 ° C (104 ° F).[83]:188 Водные млекопитающие и наоборот, они задерживают воздух в своей шерсти, чтобы сохранить тепло, сохраняя кожу сухой.[83]:162–163

Окраска
Шерсть млекопитающих окрашена по разным причинам, в том числе из основных факторов селективного давления: камуфляж, половой отбор, общение и терморегуляция. Окраска волос и кожи млекопитающих в основном определяется типом и количеством меланин; эумеланины для коричневого и черного цветов и феомеланин для диапазона от желтого до красновато-коричневого цвета, что дает млекопитающим земной тон.[103][104] Некоторые млекопитающие, такие как мандрил, имеют более яркие цвета благодаря структурная окраска.[105] Многие ленивцы кажутся зелеными, потому что их мех покрыт зеленым водоросли; это может быть симбиотический отношение, которое дает камуфляж ленивцам.[106]
Камуфляж оказывает сильное влияние на большое количество млекопитающих, поскольку помогает укрыть особей от хищников или добычи.[107] У арктических и субарктических млекопитающих, таких как Арктическая лиса (Алопекс лагопус), ошейник лемминг (Dicrostonyx groenlandicus), горностай (Mustela горностая), и заяц на снегоступах (Lepus americanus), сезонная смена цвета Цвет между коричневым летом и белым зимой в значительной степени обусловлен маскировкой.[108] Некоторые древесные млекопитающие, особенно приматы и сумчатые, имеют оттенки фиолетового, зеленого или синего цвета кожи на частях тела, что указывает на некоторые явные преимущества в их большей части. древесный среда обитания из-за конвергентная эволюция.[105]
Апосематизм, предупреждение возможных хищников, является наиболее вероятным объяснением черно-белого шкуры многих млекопитающих, которые могут защищаться, например, в зловонном скунс и мощный и агрессивный медоед.[109] Цвет шерсти иногда бывает сексуально диморфный, как в многие виды приматов.[110] Различия в окрасе шерсти самок и самцов могут указывать на уровень питания и гормонов, что важно при выборе партнера.[111] Цвет шерсти может влиять на способность сохранять тепло в зависимости от того, сколько света отражается. Млекопитающие с более темной шерстью могут поглощать больше тепла от солнечного излучения и оставаться в тепле, а некоторые более мелкие млекопитающие, такие как полевки, иметь темный мех зимой. Белый бесцветный мех арктических млекопитающих, таких как белый медведь, может отражать больше солнечного излучения непосредственно на кожу.[83]:166–167[102] Ослепительная черно-белая полоса зебры по-видимому, обеспечивают некоторую защиту от укусов мух.[112]
Репродуктивная система

В мужской плаценте пенис используется как при мочеиспускании, так и при совокуплении. В зависимости от вида эрекция может подпитываться кровотоком в сосудистую, губчатую ткань или мышечным действием. Пенис может содержаться в крайняя плоть когда не эрегированный, и некоторые плаценты также имеют кость полового члена (бакулюм ).[113] У сумчатых обычно пенисы раздвоены,[114] в то время как ехидна пенис обычно имеет четыре головки и только две функционируют.[115] В яички большинства млекопитающих спускаются в мошонка который обычно находится кзади от полового члена, но часто кпереди у сумчатых. Самки млекопитающих обычно имеют клитор, большие половые губы и малые половые губы снаружи, а внутренняя система содержит парные яйцеводы, 1-2 матка, 1-2 услуги и влагалище. У сумчатых есть два боковых влагалища и медиальное влагалище. «Влагалище» монотремов лучше понимать как «урогенитальный синус». Маточные системы плацентарных млекопитающих могут варьироваться в зависимости от дуплекса, если есть две матки и шейки, которые открываются во влагалище, двудольные, если две маточные рога имеют единственную шейку матки, которая соединяется с влагалищем, двурогую, состоящую из двух маточных рогов, которые соединены дистально, но разделены медиально, образуя Y-образную форму, и симплекс, который имеет единственную матку.[116][117][83]:220–221, 247

Предковым условием для размножения млекопитающих является рождение относительно неразвитых либо в результате прямого живорождения, либо в течение короткого периода в виде яиц с мягкой скорлупой. Вероятно, это связано с тем, что туловище не могло расширяться из-за наличия надлобковые кости. Самая старая демонстрация этого репродуктивного стиля связана с Кайентатерий, которые производили неосвоенные перинирует, но размер помета намного больше, чем у любого современного млекопитающего - 38 особей.[118] Большинство современных млекопитающих живородящий, рожая жить молодым. Однако пять видов монотрем, утконос и четыре вида ехидны, откладывают яйца. Монотремы имеют система определения пола отличается от большинства других млекопитающих.[119] В частности, половые хромосомы утконоса больше похожи на куриного, чем на терианского млекопитающего.[120]
Живородящие млекопитающие относятся к подклассу Theria; те, что живут сегодня, находятся в сумчатых и плацентарных инфраклассах. У сумчатых короткое беременность период, как правило, короче, чем эстральный цикл и рождает неразвитого новорожденного, который затем подвергается дальнейшему развитию; у многих видов это происходит в мешочковидном мешочке, сумчатый, расположенный в передней части материнского брюшная полость. Это плезиоморфный состояние среди живородящих млекопитающих; наличие надлобковых костей у всех неплацентарных млекопитающих предотвращает расширение туловища, необходимое для полной беременности.[74] Вероятно, даже бесплацентарные эвтерианцы размножались таким образом.[121] Плаценты рождают относительно полных и развитых детенышей, обычно после длительных периодов беременности.[122] Они получили свое название от плацента, который соединяет развивающийся плод со стенкой матки, обеспечивая усвоение питательных веществ.[123] У плацентарных млекопитающих надлобковая область либо полностью утрачена, либо превращается в бакулюм; позволяя туловищу расширяться, и таким образом рождается развитое потомство.[118]
В молочные железы млекопитающих специализируются на производстве молока, основного источника питания для новорожденных. Односторонние животные рано ответвились от других млекопитающих и не имеют соски наблюдается у большинства млекопитающих, но у них есть молочные железы. Молодые слизывают молоко с участка молочной железы на животе матери.[124] По сравнению с плацентарными млекопитающими молоко сумчатых сильно меняется как по продуктивности, так и по питательному составу из-за слаборазвитого молодняка. Кроме того, молочные железы обладают большей автономией, что позволяет им кормить детенышей на разных стадиях развития разными видами молока.[125] Лактоза является основным сахаром в молоке плацентарных млекопитающих, в то время как в молоке одноплодных и сумчатых животных преобладают олигосахариды.[126] Отлучение от груди это процесс, при котором млекопитающее становится менее зависимым от материнского молока и больше от твердой пищи.[127]
Эндотермия
Почти все млекопитающие эндотермический («теплокровный»). У большинства млекопитающих также есть волосы, чтобы согреться. Как и птицы, млекопитающие могут добывать корм или охотиться в погоду и климат, слишком холодный для экзотермический («хладнокровные») рептилии и насекомые. Эндотермия требует большого количества пищевой энергии, поэтому млекопитающие едят больше пищи на единицу веса, чем большинство рептилий.[128] Маленькие насекомоядные млекопитающие едят огромные количества для своего размера. Редкое исключение, голый землекоп выделяет мало тепла для обмена веществ, поэтому считается рабочим пойкилотерм.[129] Птицы также обладают эндотермией, поэтому эндотермия характерна не только для млекопитающих.[130]
Продолжительность жизни вида
Среди млекопитающих максимальная продолжительность жизни видов значительно различается (например, землеройка имеет продолжительность жизни два года, тогда как самый старый гренландский кит составляет 211 лет).[131] Хотя основная причина этих различий в продолжительности жизни все еще не ясна, многочисленные исследования показывают, что способность к восстановить повреждение ДНК является важным фактором продолжительности жизни млекопитающих. В исследовании 1974 г., проведенном Харт и Сетлоу,[132] Было обнаружено, что у семи видов млекопитающих способность к репарации путем эксцизии систематически повышалась с увеличением продолжительности жизни. Было обнаружено, что продолжительность жизни видов надежно коррелирует со способностью распознавать двухцепочечные разрывы ДНК, а также с уровнем белка репарации ДНК. Ku80.[131] При исследовании клеток шестнадцати видов млекопитающих было обнаружено, что гены, используемые в репарации ДНК, регулируемый у более долгоживущих видов.[133] Клеточный уровень фермента репарации ДНК поли-АДФ-рибоза-полимераза было обнаружено, что коррелирует с продолжительностью жизни вида в исследовании 13 видов млекопитающих.[134] Три дополнительных исследования различных видов млекопитающих также сообщили о корреляции между продолжительностью жизни вида и способностью к восстановлению ДНК.[135][136][137]
Передвижение
Наземный

Большинство позвоночных - амфибии, рептилии и некоторые млекопитающие, такие как люди и медведи - являются стопоходящий, ходьба всей нижней частью стопы. Многие млекопитающие, такие как кошки и собаки, пальцевидный при ходьбе на носках, большая длина шага обеспечивает большую скорость. Пальчатые млекопитающие также часто умеют тихо двигаться.[138] Некоторые животные, такие как лошади находятся унгулиграда, ходить на цыпочках. Это еще больше увеличивает длину их шага и, следовательно, скорость.[139] Некоторые млекопитающие, а именно человекообразные обезьяны, также известны как ходить на костяшках пальцев, по крайней мере, для их передних ног. Гигантские муравьеды[140] и утконосы[141] также ходят костяшками пальцев. Некоторые млекопитающие двуногие, используя только две конечности для передвижения, что можно увидеть, например, у людей и человекообразных обезьян. Двуногие виды имеют большее поле зрения, чем четвероногие, экономят больше энергии и обладают способностью манипулировать объектами руками, что помогает в поисках пищи. Вместо ходьбы некоторые двуногие, например, кенгуру и кенгуровые крысы.[142][143]
Животные будут использовать разные походки для разных скоростей, местности и ситуаций. Например, лошади демонстрируют четыре естественных аллюра, самый медленный походка лошади это ходить, то есть три более быстрых походки, которые, от самой медленной до самой быстрой, являются рысь, то галоп и галопом. У животных также может быть необычная походка, которую иногда используют, например, для движения вбок или назад. Например, основной человеческие походки двуногие ходьба и Бег, но иногда они используют много других походок, в том числе четвероногую ползти в ограниченном пространстве.[144] Млекопитающие показывают широкий спектр походки, порядок, в котором они размещают и поднимают свои придатки в движении. Походки можно сгруппировать по категориям в соответствии с их паттернами поддерживающей последовательности. У четвероногих есть три основные категории: ходьба, бег и походка. прыжки.[145] Ходьба - это наиболее распространенная походка, при которой некоторые ноги в любой момент времени находятся на земле, и она встречается почти у всех животных с ногами. Считается, что бег происходит, когда в некоторых точках шага все ноги отрываются от земли в момент подвешивания.[144]
Древесный

У древесных животных часто есть удлиненные конечности, которые помогают им преодолевать пропасти, добираться до фруктов или других ресурсов, проверять прочность опоры впереди и, в некоторых случаях, брахиат (качаться между деревьями).[146] Многие древесные виды, такие как древесные дикобразы, шелковистые муравьеды, паукообразные обезьяны и опоссумы, использовать цепкие хвосты хвататься за ветки. У обезьяны-паука кончик хвоста имеет либо оголенный участок, либо липкую подушечку, что обеспечивает повышенное трение. Когти можно использовать для взаимодействия с грубым субстратом и изменения направления сил, применяемых животным. Это то, что позволяет белки лазить по стволам деревьев, которые настолько велики, что с точки зрения такого маленького животного они кажутся практически плоскими. Однако когти могут мешать животному хватать очень маленькие ветки, так как они могут слишком сильно обхватить животное и уколоть лапу. Приматы используют фрикционный захват, полагаясь на безволосые кончики пальцев. Сжатие ветки между кончиками пальцев создает силу трения, которая удерживает руку животного на ветке. Однако этот тип захвата зависит от угла силы трения, то есть от диаметра ответвления, при этом более крупные ответвления приводят к снижению способности захвата. Чтобы контролировать спуск, особенно по веткам большого диаметра, некоторые древесные животные, такие как белки, развили очень подвижные голеностопные суставы, которые позволяют вращать ступню в «обратную» позу. Это позволяет когтям цепляться за шероховатую поверхность коры, противодействуя силе тяжести. Небольшой размер дает древесным видам много преимуществ: например, увеличение относительного размера ветвей по отношению к животному, более низкий центр масс, повышенная устойчивость, меньшая масса (позволяющая передвигаться по меньшим ветвям) и способность перемещаться по более загроможденной среде обитания.[146] Размер, связанный с весом, влияет на планирующих животных, таких как сахарный планер.[147] Некоторые виды приматов, летучих мышей и все виды лень добиться пассивной устойчивости, подвешиваясь под веткой. И подача, и опрокидывание становятся неуместными, поскольку единственный способ потерпеть неудачу - потерять сцепление.[146]
Антенна

Летучие мыши - единственные млекопитающие, которые действительно могут летать. Они летают по воздуху с постоянной скоростью, перемещая свои крылья вверх и вниз (обычно также с некоторым движением вперед-назад). Поскольку животное находится в движении, относительно его тела образуется некоторый воздушный поток, который в сочетании со скоростью крыльев создает более быстрый воздушный поток, движущийся над крылом. Это создает вектор подъемной силы, направленный вперед и вверх, и вектор силы сопротивления, направленный назад и вверх. Направленные вверх компоненты противодействуют силе тяжести, удерживая тело в воздухе, в то время как передний компонент обеспечивает тягу, чтобы противодействовать сопротивлению как крыла, так и корпуса в целом.[148]
Крылья летучих мышей намного тоньше и состоят из большего количества костей, чем у птиц, что позволяет летучим мышам более точно маневрировать и летать с большей подъемной силой и меньшим сопротивлением.[149][150] Сложив крылья внутрь к телу при движении вверх, они потребляют на 35% меньше энергии во время полета, чем птицы.[151] Мембраны нежные, легко рвутся; однако ткань перепонки летучей мыши способна вырасти заново, так что небольшие разрывы могут быстро зажить.[152] Поверхность их крыльев оборудована сенсорными рецепторами на небольших выступах, называемых Ячейки Меркель, также встречается на кончиках пальцев человека. Эти чувствительные области у летучих мышей различны, так как каждая неровность имеет крошечный волос в центре, что делает его еще более чувствительным и позволяет летучей мыши обнаруживать и собирать информацию о воздухе, текущем над ее крыльями, а также летать более эффективно, изменяя форму. крыльев в ответ.[153]
Ископаемые и подземные


Ископаемое (от лат. фоссор, что означает «копатель») - животное, приспособленное к рытью земли, которое живет в основном, но не только под землей. Некоторые примеры барсуки, и голый землекоп. Много грызун виды также считаются ископаемыми, поскольку большую часть дня, но не весь день, живут в норах. Виды, которые живут исключительно под землей, являются подземными, а виды с ограниченной адаптацией к окаменелому образу жизни - подземными. Некоторые организмы окаменелости, чтобы помочь в регулирование температуры в то время как другие используют подземную среду обитания для защиты от хищники или для хранение продуктов.[154]
Ископаемые млекопитающие имеют веретеновидное тело, наиболее толстое в плечах и сужающееся к хвосту и носу. Неспособность видеть в темных норах, у большинства из них дегенерированные глаза, но дегенерация варьируется между видами; карманные суслики, например, являются только полуфоссориальными и имеют очень маленькие, но функциональные глаза в полностью ископаемых сумчатый крот глаза выродились и бесполезны, тальпа родинки имеют рудиментарный глаза и мыс золотой крот имеет слой кожи, покрывающий глаза. Наружные уши тоже очень маленькие или отсутствуют. У настоящих окаменелых млекопитающих короткие и крепкие ноги, поскольку для роющего млекопитающего сила важнее скорости, но у полуфоссориальных млекопитающих есть беглый ноги. Передние лапы широкие и имеют сильные когти, которые помогают разрыхлить грязь при рытье нор, а задние лапы имеют перепонки и когти, которые помогают отбрасывать разрыхленную землю назад. У большинства есть большие резцы, чтобы грязь не попала в рот.[155]
Многие ископаемые млекопитающие, такие как землеройки, ежи и кроты, были классифицированы в соответствии с ныне устаревшим порядком. Насекомоядные.[156]
Водный

Полностью водные млекопитающие, китообразные и сирены, потеряли ноги и имеют хвостовой плавник, чтобы двигаться по воде. Движение флиппера непрерывное. Киты плавают, перемещая хвостовой плавник и нижнюю часть тела вверх и вниз, двигаясь вертикально, в то время как их ласты в основном используются для управления. Их анатомия скелета позволяет им быстро плавать. Большинство видов имеют спинной плавник чтобы они не перевернулись в воде.[157][158] Трематоды сирен поднимаются вверх и вниз длинными движениями, чтобы продвинуть животное вперед, и их можно повернуть, чтобы повернуть. Передние конечности - это ласты, похожие на весло, которые помогают в поворотах и замедлении.[159]
Полуводный у млекопитающих, как и у ластоногих, есть две пары плавников на передней и задней части, передние и задние. Локти и лодыжки прижаты к телу.[160][161] У ластоногих есть несколько приспособлений для уменьшения тащить. Помимо обтекаемых тел, у них есть гладкая сеть мышечные пучки в их коже, которая может увеличиваться ламинарный поток и облегчить им скольжение в воде. Им также не хватает арректор пили, поэтому их мех может быть гладким во время плавания.[162] Они полагаются на свои передние ласты для передвижения подобно крыльям. пингвины и морские черепахи.[163] Движение передних ласт непостоянно, и животное скользит между каждым гребком.[161] По сравнению с наземными хищниками длина передних конечностей уменьшена, что дает локомоторным мышцам плечевого и локтевого суставов большее механическое преимущество;[160] задние ласты служат стабилизаторами.[162] К другим полуводным млекопитающим относятся бобры, бегемоты, выдры и утконосы.[164] Бегемоты - очень крупные полуводные млекопитающие, и их бочкообразные тела имеют гравипортальный скелетные структуры,[165] адаптированы к их огромному весу, а их удельный вес позволяет им тонуть и двигаться по дну реки.[166]
Поведение
Общение и вокализация

Многие млекопитающие общаются голосом. Голосовое общение служит многим целям, в том числе в брачных ритуалах, например предупреждающие звонки,[168] для обозначения источников питания и в социальных целях. Самцы часто звонят во время брачных ритуалов, чтобы отогнать других самцов и привлечь самок, как в рев из львы и красный олень.[169] В песни горбатого кита могут быть сигналами самкам;[170] у них разные диалекты в разных регионах океана.[171] Социальные вокализации включают территориальный звонки гиббоны, и использование частоты в большие копьеносые летучие мыши различать группы.[172] В верветка обезьяна подает отчетливый сигнал тревоги для каждого из по крайней мере четырех разных хищников, а реакция других обезьян варьируется в зависимости от этого сигнала. Например, если сигнал тревоги сигнализирует питону, обезьяны забираются на деревья, тогда как сигнал тревоги орла заставляет обезьян искать укрытие на земле.[167] Луговые собачки аналогично имеют сложные сигналы, которые сигнализируют о типе, размере и скорости приближающегося хищника.[173] Слоны общаются в обществе с помощью различных звуков, включая фырканье, крик, трубку, рев и урчание. Некоторые из громких звонков инфразвуковой, ниже диапазона слышимости людей, и могут быть услышаны другими слонами на расстоянии до 6 миль (9,7 км) в неподвижное время около восхода и заката.[174]
Млекопитающие передают сигнал разными способами. Многие дают визуальные сигналы против хищников, как когда олень и газель Stot, честно указывающий их пригодное состояние и их способность к побегу,[175][176] или когда белохвостый олень и другие хищные млекопитающие при тревоге выделяют заметные отметины на хвосте, информируя хищника о том, что он был обнаружен.[177] Многие млекопитающие используют ароматическая маркировка, иногда, возможно, для защиты территории, но, вероятно, с рядом функций как внутри, так и между видами.[178][179][180] Микробаты и зубатые киты включая океанические дельфины озвучивать как социально, так и в эхолокация.[181][182][183]
Кормление

Поддержание постоянной высокой температуры тела требует больших затрат энергии, поэтому млекопитающим требуется обильный и питательный рацион. Хотя самые ранние млекопитающие, вероятно, были хищниками, с тех пор различные виды адаптировались для удовлетворения своих диетических потребностей различными способами. Некоторые едят других животных - это плотоядный диета (и включает в себя диеты на насекомоядных). Другие млекопитающие, называемые травоядные животные, ешьте растения, содержащие сложные углеводы например, целлюлоза. Диета для травоядных включает такие подтипы, как зерно (поедание семян), фоливия (поедание листьев), плодоядный (есть фрукты), нектароядный (поедание нектара), жевательная резинка (поедание жевательной резинки) и микофагия (поедание грибка). Пищеварительный тракт травоядного животного является хозяином бактерий, которые ферментируют эти сложные вещества и делают их доступными для пищеварения, которые либо находятся в многокамерном корпусе. желудок или в большой слепой кишке.[87] Некоторые млекопитающие копрофаг, потребляя кал для поглощения питательных веществ, которые не были усвоены при первом приеме пищи.[83]:131–137 An всеядный ест и добычу, и растения. У хищных млекопитающих есть простой пищеварительный тракт поскольку белки, липиды и минералы содержащиеся в мясе, не требуют специального пищеварения. Исключения из этого включают усатые киты кто также дом Кишечная флора в многокамерном желудке, как у наземных травоядных.[184]
Размер животного также является фактором при определении типа диеты (Правило Аллена ). Поскольку у мелких млекопитающих высокое отношение площади поверхности, отводящей тепло, к объему, выделяющему тепло, они, как правило, имеют высокие потребности в энергии и скорость метаболизма. Млекопитающие, которые весят менее примерно 18 унций (510 г; 1,1 фунта), в основном являются насекомоядными, поскольку они не переносят медленный и сложный процесс пищеварения травоядных животных. С другой стороны, более крупные животные выделяют больше тепла, и теряется меньше этого тепла. Поэтому они могут переносить либо более медленный процесс сбора (плотоядные, питающиеся более крупными позвоночными), либо более медленный процесс пищеварения (травоядные).[185] Кроме того, млекопитающие, которые весят более 18 унций (510 г; 1,1 фунта), обычно не могут собрать достаточно насекомых в часы бодрствования, чтобы выжить. Единственные крупные насекомоядные млекопитающие - это те, которые питаются огромными колониями насекомых (муравьи или же термиты ).[186]


Некоторые млекопитающие всеядны и демонстрируют разную степень хищничества и травоядности, обычно склоняясь к одному больше, чем к другому. Поскольку растения и мясо перевариваются по-разному, предпочтение отдается одному перед другим, как, например, у медведей, у которых одни виды могут быть в основном плотоядными, а другие - в основном травоядными.[188] Они сгруппированы в три категории: мезокармоявление (50–70% мяса), гиперплетение (70% и более мяса), и лицемерие (50% или менее мяса). Зубной ряд гипокарноядных состоит из тупых треугольных хищных зубов, предназначенных для измельчения пищи. Однако у гиперплотоядных есть конические зубы и острые хищники, предназначенные для рубящих ударов, а в некоторых случаях - сильные челюсти для дробления костей, как в случае гиены, позволяя им поедать кости; некоторые вымершие группы, особенно Machairodontinae, имел саблевидный клыки.[187]
Некоторые физиологические плотоядные животные потребляют растительные вещества, а некоторые физиологические травоядные - мясо. С точки зрения поведения это сделало бы их всеядными, но с физиологической точки зрения это может быть связано с зоофармакогнозия. С физиологической точки зрения животные должны иметь возможность получать энергию и питательные вещества из растительных и животных материалов, чтобы считаться всеядными. Таким образом, таких животных все еще можно классифицировать как плотоядных и травоядных, если они просто получают питательные вещества из материалов, происходящих из источников, которые, казалось бы, не дополняют их классификацию.[189] Например, хорошо известно, что некоторые копытные, такие как жирафы, верблюды и крупный рогатый скот, грызут кости, чтобы потреблять определенные минералы и питательные вещества.[190] Кроме того, кошки, которых обычно считают облигатными плотоядными животными, иногда едят траву, чтобы извергнуть неперевариваемый материал (например, комки волос ), способствует выработке гемоглобина и действует как слабительное.[191]
Многие млекопитающие в отсутствие достаточных потребностей в пище в окружающей среде подавляют свой метаболизм и сохраняют энергию в процессе, известном как спячка.[192] В период, предшествующий зимней спячке, более крупные млекопитающие, такие как медведи, становятся полифагический чтобы увеличить запасы жира, тогда как более мелкие млекопитающие предпочитают собирать и прятать пищу.[193] Замедление метаболизма сопровождается снижением частоты сердечных сокращений и дыхания, а также падением внутренней температуры, которая в некоторых случаях может быть близка к температуре окружающей среды. Например, внутренняя температура гибернации арктические суслики может опускаться до -2,9 ° C (26,8 ° F), однако голова и шея всегда остаются выше 0 ° C (32 ° F).[194] Несколько млекопитающих в жарких условиях летучий во время засухи или сильной жары, например толстохвостый карликовый лемур (Cheirogaleus medius).[195]
Интеллект
У разумных млекопитающих, таких как приматы, то головной мозг больше по сравнению с остальной частью мозга. Интеллект Сам по себе не просто определить, но признаки интеллекта включают способность к обучению в сочетании с поведенческой гибкостью. Крысы, например, считаются очень умными, так как они могут учиться и выполнять новые задачи, способность, которая может быть важна, когда они впервые заселяют свежий среда обитания. У некоторых млекопитающих сбор пищи, по-видимому, связан с интеллектом: у оленя, питающегося растениями, мозг меньше, чем у кошки, которая должна думать, чтобы перехитрить свою добычу.[186]

Использование инструмента животными может указывать на разные уровни учусь и познание. В морская выдра использует камни как неотъемлемую и постоянную часть своего кормового поведения (разбивание морское ушко из камней или вскрытия ракушек), при этом некоторые народы тратят 21% своего времени на изготовление инструментов.[196] Использование других инструментов, например шимпанзе использование веток для «ловли» термитов, может быть развито наблюдая, как другие используют инструменты и может даже быть настоящим примером обучения животных.[197] Инструменты могут даже использоваться для решения головоломок, в которых животное, кажется, испытывает "Момент Эврики".[198] Другие млекопитающие, не использующие инструменты, например собаки, также могут испытать момент Эврики.[199]
Размер мозга Ранее считалось основным показателем интеллекта животного. Поскольку большая часть мозга используется для поддержания функций организма, большее соотношение мозг к массе тела может увеличить объем мозговой массы, доступной для более сложных когнитивных задач. Аллометрический анализ показывает, что размер мозга млекопитающих составляет примерно2⁄3 или3⁄4 показатель массы тела. Сравнение размера мозга конкретного животного с ожидаемым размером мозга на основе такого аллометрического анализа дает коэффициент энцефализации это можно использовать как еще один показатель интеллекта животных.[200] Кашалоты имеют самую большую массу мозга среди всех животных на земле, в среднем 8000 кубических сантиметров (490 дюймов).3) и 7,8 кг (17 фунтов) у зрелых самцов.[201]
Самосознание кажется признаком абстрактного мышления. Считается, что самосознание, хотя и недостаточно четко определено, является предшественником более продвинутых процессов, таких как метакогнитивное мышление. Традиционный метод измерения - это зеркальный тест, который определяет, обладает ли животное способностью к самопознанию.[202] Млекопитающие, прошедшие тест на зеркало, включают азиатских слонов (некоторые проходят, некоторые нет);[203] шимпанзе;[204] бонобо;[205] орангутаны;[206] люди, от 18 месяцев (зеркальная сцена );[207] афалины[а][208] косатки;[209] и ложных косаток.[209]
Социальная структура


Эусоциальность это высший уровень социальной организации. В этих обществах пересекаются взрослые поколения, есть разделение репродуктивного труда и совместная забота о молодежи. Обычно насекомые, такие как пчелы, муравьи и термиты, обладают эусоциальным поведением, но оно проявляется у двух видов грызунов: голого землекопа.[210] и Дамаралендский землекоп.[211]
Досоциальность - это когда животные демонстрируют больше, чем просто сексуальные взаимодействия с представителями одного и того же вида, но не могут считаться эусоциальными. То есть досоциальные животные могут демонстрировать совместную жизнь, совместную заботу о молодняке или примитивное разделение репродуктивного труда, но они не проявляют всех трех основных черт эусоциальных животных. Люди и некоторые виды Callitrichidae (мартышки и тамарины ) уникальны среди приматов по степени совместной заботы о молодняке.[212] Гарри Харлоу поставить эксперимент с макаки резус, досоциальные приматы - в 1958 г .; Результаты этого исследования показали, что социальные контакты необходимы молодым обезьянам для умственного и сексуального развития.[213]
А общество ядерного синтеза это общество, которое часто меняется по своим размерам и составу, образуя постоянную социальную группу, называемую «родительской группой». Постоянные социальные сети состоят из всех отдельных членов сообщества и часто меняются, чтобы отслеживать изменения в их среде. В обществе деления-слияния основная родительская группа может распадаться (делиться) на более мелкие стабильные подгруппы или индивидов, чтобы адаптироваться к относящийся к окружающей среде или социальные обстоятельства. Например, некоторые самцы могут оторваться от основной группы, чтобы охотиться или добывать пищу в течение дня, но ночью они могут вернуться, чтобы присоединиться (слиться) с основной группой, чтобы разделить пищу и принять участие в других занятиях. Это проявляют многие млекопитающие, например приматы (например, орангутаны и паучьи обезьяны ),[214] слоны[215] пятнистые гиены,[216] львы[217] и дельфины.[218]
Одиночные животные защищают территорию и избегают социальных взаимодействий с представителями своего вида, за исключением периода размножения. Это сделано для того, чтобы избежать конкуренции за ресурсы, поскольку две особи одного вида занимают одну и ту же нишу, а также для предотвращения истощения запасов пищи.[219] Одиночное животное, добывая пищу, может быть менее заметным для хищников или добычи.[220]

В иерархия, люди либо доминируют, либо подчиняются. Деспотическая иерархия - это когда один человек доминирует, а другие подчиняются, как у волков и лемуров.[221] и иерархия представляет собой линейный рейтинг индивидов, в котором есть высший и нижний индивиды. Клевета также может быть ранжирована по полу, где низший представитель пола имеет более высокий рейтинг, чем высший представитель другого пола, как у гиен.[222] Доминирующие особи или альфы имеют высокий шанс на репродуктивный успех, особенно в гаремы где один или несколько самцов (постоянные самцы) имеют исключительные права на размножение самок в группе.[223] Самцы-нерезиденты также могут быть приняты в гаремы, но некоторые виды, такие как обычная летучая мышь-вампир (Desmodus rotundus), может быть более строгим.[224]
Некоторые млекопитающие прекрасно моногамный, что означает, что они помощник на всю жизнь и не берите других партнеров (даже после смерти первого супруга), как с волками, Евразийские бобры, и выдры.[225][226] Существует три типа полигамии: один или несколько доминирующих самцов имеют право на размножение (многоженство ), несколько мужчин, с которыми спариваются женщины (полиандрия), или несколько мужчин имеют исключительные отношения с несколькими женщинами (полиандрия). Гораздо чаще случаются полигинные спаривания, которые, за исключением токи, по оценкам, встречаются у 90% млекопитающих.[227] Спаривание лек происходит, когда самцы собираются вокруг самок и пытаются привлечь их различными демонстрации ухаживания и вокализации, как у тюленей.[228]
Все высшие млекопитающие (за исключением монотремов) разделяют две основные адаптации для ухода за детенышами: живорождение и лактация. Это подразумевает возможность выбора степени родительская забота. Они могут строить гнезда и рыть норы, чтобы вырастить своих детенышей, или кормить и охранять их часто в течение длительного периода времени. Многие млекопитающие K-selected, и вкладывают в своих детей больше времени и энергии, чем r-выбранный животные. Когда два животных спариваются, они оба заинтересованы в успехе потомства, хотя часто в разной степени. Самки млекопитающих проявляют некоторую степень материнской агрессии, что является еще одним примером родительской заботы, которая может быть направлена против других самок этого вида или детенышей других самок; однако некоторые млекопитающие могут «воспитывать» детенышей других самок и заботиться о них. Самцы млекопитающих могут играть определенную роль в воспитании детей, как и тенреки, однако это варьируется от вида к виду, даже в пределах одного рода. Например, самцы южная коснохвостая макака (Макака неместрина) не участвуют в уходе за детьми, в то время как мужчины Японская макака (М. fuscata) делать.[229]
Люди и другие млекопитающие
В человеческой культуре

Нечеловеческие млекопитающие играют в человеческой культуре самые разные роли. Они самые популярные из домашние питомцы, с десятками миллионов собак, кошек и других животных, включая кролики и мышей, которых содержат семьи по всему миру.[230][231][232] Млекопитающие, такие как мамонты, лошади и олени являются одними из самых ранних предметов искусства, найденных в Верхний палеолит наскальные рисунки например, в Ласко.[233] Крупные художники, такие как Альбрехт Дюрер, Джордж Стаббс и Эдвин Ландсир известны своими портретами млекопитающих.[234] Многие виды млекопитающих были охотился для спорта и для еды; олень и дикий кабан особенно популярны как игровые животные.[235][236][237] Млекопитающие, такие как лошади и собаки широко участвуют в спортивных состязаниях, часто в сочетании с ставка на исход.[238][239] Существует противоречие между ролью животных как товарищей по отношению к людям и их существованием как индивидов с собственные права.[240] Млекопитающие также играют самые разные роли в литературе,[241][242][243] фильм[244] мифология и религия.[245][246][247]
Использование и важность

Одомашненный млекопитающие составляют большую часть домашний скот поднял для мясо по всему миру. В их числе (2009 г.) около 1,4 млрд. крупный рогатый скот, 1000000000 овца, 1000000000 домашние свиньи,[248][249] и (1985) более 700 миллионов кроликов.[250] Рабочие домашние животные в том числе крупный рогатый скот и лошади использовались для работы и транспорт от истоков сельского хозяйства, их численность сокращается с появлением механизированного транспорта и сельскохозяйственная техника. В 2004 году они по-прежнему обеспечивали около 80% электроэнергии в основном небольшими фермами в странах третьего мира и около 20% мирового транспорта, опять же в основном в сельской местности. В горных районах, непригодных для колесной техники, вьючные животные продолжать перевозить грузы.[251]Шкуры млекопитающих обеспечивают натуральная кожа за туфли, одежда и обивка.[252] Шерсть от млекопитающих, включая овец, коз и альпаки веками использовался для изготовления одежды.[253][254] Млекопитающие играют важную роль в науке, поскольку экспериментальные животные как в фундаментальных биологических исследованиях, например, в генетике,[255] и в разработке новых лекарств, которые должны быть тщательно протестированы, чтобы продемонстрировать их безопасность.[256] Миллионы млекопитающих, особенно мышей и крыс, используются в эксперименты каждый год.[257] А нокаутирующая мышь это генетически модифицированная мышь с инактивированным ген, заменены или повреждены искусственным фрагментом ДНК. Они позволяют изучать последовательный гены, функции которых неизвестны.[258] Небольшой процент млекопитающих - нечеловеческие приматы, которые используются в исследованиях из-за их сходства с людьми.[259][260][261]
Чарльз Дарвин, Джаред Даймонд и другие отметили важность домашних млекопитающих в Неолитическое развитие сельского хозяйства и из цивилизация, заставляя фермеров заменять охотники-собиратели во всем мире.[b][263] Этот переход от охоты и собирательства к выпас стада и растущий урожай был важным шагом в истории человечества. Новые сельскохозяйственные экономики, основанные на одомашненных млекопитающих, вызвали «радикальную реструктуризацию человеческих обществ, глобальные изменения в биоразнообразии и значительные изменения в формах суши и атмосферы Земли ... важные результаты».[264]
Гибриды


Гибриды - это потомство, полученное в результате скрещивания двух генетически разных особей, что обычно приводит к высокой степени гетерозиготности, хотя гибрид и гетерозиготность не являются синонимами. Преднамеренное или случайное скрещивание двух или более видов близкородственных животных посредством разведения в неволе - это деятельность человека, которая существует на протяжении тысячелетий и росла в экономических целях.[265] Гибриды между разными подвидами внутри вида (например, между бенгальский тигр и Сибирский тигр ) известны как внутривидовые гибриды. Гибриды между разными видами одного и того же рода (например, между львами и тиграми) известны как межвидовые гибриды или кроссы. Гибриды между разными родами (например, между овцами и козами) известны как межродовые гибриды.[266] Естественные гибриды произойдут в гибридные зоны, где две популяции видов одного и того же рода или вида, живущие в одной или соседней местности, будут скрещиваться друг с другом. Некоторые гибриды были признаны видами, например, красный волк (Хотя это спорно).[267]
Искусственный отбор преднамеренное селекция домашних животных, используется для размножаться недавно вымерший животных в попытке создать породу животных с фенотип что напоминает этот вымерший дикого типа предок. Воспроизводимый (внутривидовой) гибрид может быть очень похож на вымерший дикий тип по внешнему виду, экологической нише и в некоторой степени по генетике, но исходный Генофонд этого дикого типа потеряны навсегда с его вымирание. В результате выведенные породы в лучшем случае являются неопределенными двойниками вымерших диких видов, поскольку Черт побери относятся к зубр.[268]
Чистокровный дикие виды, эволюционировавшие в определенную экологию, могут оказаться под угрозой исчезновения[269] через процесс генетическое загрязнение, неконтролируемая гибридизация, интрогрессия генетическое заболачивание, которое приводит к гомогенизации или вне конкуренции от гетерозийный гибридный вид.[270] Когда новые популяции импортируются или селективно разводятся людьми, или когда изменение среды обитания приводит к контакту ранее изолированных видов, возможно исчезновение некоторых видов, особенно редких разновидностей.[271] Скрещивание могут затопить более редкий генофонд и создавать гибриды, истощая чистокровный генофонд. Например, исчезающие дикий буйвол находится под угрозой исчезновения из-за генетического загрязнения домашний буйвол. Такие вымирания не всегда очевидны из морфологический точка зрения. Некоторая степень поток генов это нормальный эволюционный процесс, тем не менее, гибридизация угрожает существованию редких видов.[272][273]
Угрозы

Исчезновение видов из экологических сообществ, выход из строя, в первую очередь обусловлено деятельностью человека.[274] Это привело к пустые леса, экологические сообщества обеднены крупными позвоночными животными.[275][276] в Событие четвертичного вымирания, массовое отмирание мегафауна разнообразие совпало с появлением людей, предполагая влияние человека. Одна из гипотез состоит в том, что люди охотились на крупных млекопитающих, таких как шерстистый мамонт, в исчезновение.[277][278] 2019 год Отчет о глобальной оценке биоразнообразия и экосистемных услуг к IPBES заявляет, что общая биомасса диких млекопитающих сократилось на 82 процента с начала человеческой цивилизации.[279][280] Дикие животные составляют всего 4% млекопитающих биомасса на Земле, а люди и их домашние животные составляют 96%.[281]
Прогнозируется, что различные виды вымрут в ближайшем будущем,[282] среди них носорог,[283] приматы,[284] панголины,[285] и жирафы.[286] По версии WWF 2020 Живая Планета Отчет, позвоночное животное дикая природа население сократилось на 68% с 1970 года в результате деятельности человека, особенно чрезмерное потребление, рост населения и интенсивное земледелие, что свидетельствует о том, что люди вызвали шестое массовое вымирание мероприятие.[287][288] Одна только охота угрожает сотням видов млекопитающих по всему миру.[289][290] Ученые утверждают, что растущий спрос на мясо вносит свой вклад в потеря биоразнообразия поскольку это важный фактор вырубка леса и разрушение среды обитания; богатые видами среды обитания, такие как значительная часть Тропический лес Амазонки, превращаются в сельскохозяйственные угодья для производства мяса.[291][292][293] Другое влияние - чрезмерная охота и браконьерство, что может сократить общую популяцию охотничьих животных,[294] особенно те, которые расположены недалеко от деревень,[295] как в случае с пекари.[296] Последствия браконьерства особенно заметны в торговля слоновой костью с африканскими слонами.[нужна цитата ] Морские млекопитающие подвергаются риску запутывания в рыболовных снастях, особенно китообразные, со смертностью отбросов от 65 000 до 86 000 особей ежегодно.[297]
Внимание уделяется исчезающим видам во всем мире, в частности из-за Конвенция о биологическом разнообразии, иначе известное как Рио-де-Жанейрское соглашение, которое включает 189 подписавших его стран, которые сосредоточены на выявлении исчезающих видов и местообитаний.[298] Еще одна известная природоохранная организация - МСОП, в который входят более 1200 правительственных и негосударственный организации.[299]
Недавние вымирания можно напрямую отнести к человеческому влиянию.[300][274] МСОП характеризует «недавнее» вымирание как те, которые произошли после пороговой точки 1500,[301] и около 80 видов млекопитающих вымерли с того времени и с 2015 года.[302] Некоторые виды, такие как Олень отца Давида[303] находятся вымершие в дикой природе, и выживают исключительно в неволе. Другие виды, такие как Флоридская пантера, находятся экологически вымерший, выживают в таком небольшом количестве, что практически не влияют на экосистему.[304]:318 Остальные популяции только локально вымерший (искоренен), все еще существующий в других местах, но сокращенный в распространении,[304]:75–77 как с исчезновением серые киты в Атлантический.[305]
Примечания
- ^ Уменьшение времени ожидания для приближения к зеркалу, повторяющиеся кружения головы и близкий просмотр отмеченных областей считались признаками самопознания, поскольку у них нет рук и они не могут касаться отмеченных областей.[208]
- ^ Даймонд обсудил этот вопрос в своей книге 1997 года. Оружие, микробы и сталь.[262]
Смотрите также
- Список родов млекопитающих - живые млекопитающие
- Список маммологов
- Список одноцветных и сумчатых
- Список плацентарных млекопитающих
- Список доисторических млекопитающих
- Список находящихся под угрозой исчезновения млекопитающих США
- Списки млекопитающих по размеру популяции
- Списки млекопитающих по регионам
- Млекопитающие, описанные в 2000-х годах
- Млекопитающие в культуре
Рекомендации
- ^ Воан Т.А., Райан Дж. М., Чаплевски Н.Дж. (2013). «Классификация млекопитающих». Маммология (6 изд.). Джонс и Бартлетт Обучение. ISBN 978-1-284-03209-3.
- ^ а б Салай Ф.С. (1999). «Классификация млекопитающих выше видового уровня: обзор». Журнал палеонтологии позвоночных. 19 (1): 191–195. Дои:10.1080/02724634.1999.10011133. JSTOR 4523980.
- ^ а б Уилсон Д., Ридер Д., ред. (2005). «Предисловие и вводный материал». Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джона Хопкинса. п. xxvi. ISBN 978-0-8018-8221-0. OCLC 62265494.
- ^ "Млекопитающие". Красный список видов, находящихся под угрозой исчезновения МСОП. МСОП. Апрель 2010 г.. Получено 23 августа 2016.
- ^ Бургин CJ, Colella JP, Кан П.Л., Upham NS (1 февраля 2018 г.). "Сколько существует видов млекопитающих?". Журнал маммологии. 99 (1): 1–14. Дои:10.1093 / jmammal / gyx147.
- ^ Роу Т. (1988). «Определение, диагноз и происхождение млекопитающих» (PDF). Журнал палеонтологии позвоночных. 8 (3): 241–264. Дои:10.1080/02724634.1988.10011708.
- ^ Lyell C (1871). Студенческие элементы геологии. Лондон: Джон Мюррей. п. 347. ISBN 978-1-345-18248-4.
- ^ Чифелли Р.Л., Дэвис Б.М. (декабрь 2003 г.). «Палеонтология. Сумчатое происхождение». Наука. 302 (5652): 1899–900. Дои:10.1126 / science.1092272. PMID 14671280.
- ^ Кемп Т.С. (2005). Происхождение и эволюция млекопитающих (PDF). Соединенное Королевство: Издательство Оксфордского университета. п. 3. ISBN 978-0-19-850760-4. OCLC 232311794.
- ^ Датта ПМ (2005). «Самое раннее млекопитающее с расширенным в поперечном направлении верхним коренным зубом из позднетриасовой (карнийской) формации Тики, бассейн Южного Рева Гондвана, Индия». Журнал палеонтологии позвоночных. 25 (1): 200–207. Дои:10.1671 / 0272-4634 (2005) 025 [0200: EMWTEU] 2.0.CO; 2.
- ^ Ло З, Мартин Т. (2007). "Анализ молярной структуры и филогении рода докодонтов" (PDF). Бюллетень Музея естественной истории Карнеги. 39: 27–47. Дои:10.2992 / 0145-9058 (2007) 39 [27: AOMSAP] 2.0.CO; 2. Архивировано из оригинал (PDF) 3 марта 2016 г.. Получено 8 апреля, 2013.
- ^ Маккенна М.С., Белл С.Г. (1997). Классификация млекопитающих выше видового уровня. Нью-Йорк: издательство Колумбийского университета. ISBN 978-0-231-11013-6. OCLC 37345734.
- ^ Нильссон М.А., Чураков Г., Соммер М., Тран Н.В., Земанн А., Брозиус Дж., Шмитц Дж. (Июль 2010 г.). «Отслеживание эволюции сумчатых с использованием архаичных геномных вставок ретропозонов». PLOS Биология. 8 (7): e1000436. Дои:10.1371 / journal.pbio.1000436. ЧВК 2910653. PMID 20668664.
- ^ Кригс Дж.О., Чураков Г., Кифманн М., Джордан У., Брозиус Дж., Шмитц Дж. (Апрель 2006 г.). «Ретропозиционные элементы как архив эволюционной истории плацентарных млекопитающих». PLOS Биология. 4 (4): e91. Дои:10.1371 / journal.pbio.0040091. ЧВК 1395351. PMID 16515367.
- ^ а б Нишихара Х., Маруяма С., Окада Н. (март 2009 г.). «Анализ ретропозонов и недавние геологические данные предполагают почти одновременное расхождение трех надотрядов млекопитающих». Труды Национальной академии наук Соединенных Штатов Америки. 106 (13): 5235–40. Bibcode:2009ПНАС..106.5235Н. Дои:10.1073 / pnas.0809297106. ЧВК 2655268. PMID 19286970.
- ^ Спрингер М.С., Мерфи В.Дж., Эйзирик Э., О'Брайен С.Дж. (февраль 2003 г.). «Диверсификация плацентарных млекопитающих и граница мелового и третичного периода». Труды Национальной академии наук Соединенных Штатов Америки. 100 (3): 1056–61. Bibcode:2003ПНАС..100.1056С. Дои:10.1073 / пнас.0334222100. ЧВК 298725. PMID 12552136.
- ^ Тарвер Дж. Э., Дос Рейс М., Мирараб С., Моран Р. Дж., Паркер С., О'Рейли Дж. Э. и др. (Январь 2016 г.). «Взаимоотношения плацентарных млекопитающих и пределы филогенетического вывода». Геномная биология и эволюция. 8 (2): 330–44. Дои:10.1093 / gbe / evv261. HDL:1983 / 64d6e437-3320-480d-a16c-2e5b2e6b61d4. ЧВК 4779606. PMID 26733575.
- ^ а б c Springer MS, Meredith RW, Janecka JE, Murphy WJ (сентябрь 2011 г.). «Историческая биогеография млекопитающих». Философские труды Лондонского королевского общества B. 366 (1577): 2478–502. Дои:10.1098 / rstb.2011.0023. ЧВК 3138613. PMID 21807730.
- ^ Мэн Дж, Ван И, Ли Ц (апрель 2011 г.). «Переходное среднее ухо млекопитающих из нового мелового эвтриконодонта Jehol». Природа. 472 (7342): 181–5. Bibcode:2011Натура.472..181M. Дои:10.1038 / природа09921. PMID 21490668. S2CID 4428972.
- ^ Альберг П.Е., Милнер А.Р. (апрель 1994 г.). «Происхождение и ранняя диверсификация четвероногих». Природа. 368 (6471): 507–514. Bibcode:1994Натура.368..507А. Дои:10.1038 / 368507a0. S2CID 4369342.
- ^ «Амниота - Палеос». Архивировано из оригинал на 2010-12-20.
- ^ «Обзор Synapsida - Palaeos». Архивировано из оригинал на 2010-12-20.
- ^ а б c Кемп Т.С. (июль 2006 г.). «Происхождение и раннее облучение терапевсидных рептилий, подобных млекопитающим: палеобиологическая гипотеза» (PDF). Журнал эволюционной биологии. 19 (4): 1231–47. Дои:10.1111 / j.1420-9101.2005.01076.x. PMID 16780524.
- ^ а б Беннетт А.Ф., Рубен Дж.А. (1986). «Метаболический и терморегуляторный статус терапсидов». В Hotton III N, MacLean JJ, Roth J, Roth EC (ред.). Экология и биология млекопитающих рептилий. Вашингтон, округ Колумбия: Пресса Смитсоновского института. С. 207–218. ISBN 978-0-87474-524-5.
- ^ Кермак Д.М., Кермак К.А. (1984). Эволюция характеров млекопитающих. Вашингтон, округ Колумбия: Крум Хелм. ISBN 978-0-7099-1534-8. OCLC 10710687.
- ^ Таннер Л. Х., Лукас С. Г., Чепмен М. Г. (2004). «Оценка записей и причин позднетриасовых вымираний» (PDF). Обзоры наук о Земле. 65 (1–2): 103–139. Bibcode:2004ESRv ... 65..103T. Дои:10.1016 / S0012-8252 (03) 00082-5. Архивировано 25 октября 2007 года.CS1 maint: неподходящий URL (связь)
- ^ Brusatte SL, Benton MJ, Ruta M, Lloyd GT (сентябрь 2008 г.). «Превосходство, конкуренция и оппортунизм в эволюционной радиации динозавров» (PDF). Наука. 321 (5895): 1485–8. Bibcode:2008Научный ... 321.1485B. Дои:10.1126 / science.1161833. PMID 18787166. S2CID 13393888.
- ^ Готье JA (1986). «Заурисхиевая монофилия и происхождение птиц». В Padian K (ред.). Происхождение птиц и эволюция полета. Мемуары Калифорнийской академии наук. 8. Сан-Франциско: Калифорнийская академия наук. С. 1–55.
- ^ Sereno PC (1991). «Базальные архозавры: филогенетические отношения и функциональные последствия». Воспоминания Общества палеонтологии позвоночных. 2: 1–53. Дои:10.2307/3889336. JSTOR 3889336.
- ^ MacLeod N, Rawson PF, Forey PL, Banner FT, Boudagher-Fadel MK, Bown PR и др. (1997). «Меловой – третичный биотический переход». Журнал геологического общества. 154 (2): 265–292. Bibcode:1997JGSoc.154..265M. Дои:10.1144 / gsjgs.154.2.0265. S2CID 129654916.
- ^ Хант Д.М., Хэнкинс М.В., Коллин С.П., Маршалл, штат Нью-Джерси. Эволюция визуальных и невизуальных пигментов. Лондон: Спрингер. п. 73. ISBN 978-1-4614-4354-4. OCLC 892735337.
- ^ Бакалар Н (2006). Найден «юрский бобер»; переписывает историю млекопитающих ». Новости National Geographic. Получено 28 мая 2016.
- ^ Холл MI, Камилар JM, Кирк EC (декабрь 2012 г.). «Форма глаз и ночное узкое место млекопитающих». Труды Королевского общества B: биологические науки. 279 (1749): 4962–8. Дои:10.1098 / rspb.2012.2258. ЧВК 3497252. PMID 23097513.
- ^ Луо ZX (декабрь 2007 г.). «Трансформация и диверсификация в ранней эволюции млекопитающих». Природа. 450 (7172): 1011–9. Bibcode:2007 Натур.450.1011L. Дои:10.1038 / природа06277. PMID 18075580. S2CID 4317817.
- ^ Пикрелл Дж. (2003). «Древнейшее ископаемое сумчатое животное, найденное в Китае». Новости National Geographic. Получено 28 мая 2016.
- ^ а б Луо ZX, Юань CX, Мэн QJ, Цзи Q (август 2011). «Юрское эвтериальное млекопитающее и расхождение сумчатых и плацентарных». Природа. 476 (7361): 442–5. Bibcode:2011Натура.476..442L. Дои:10.1038 / природа10291. PMID 21866158. S2CID 205225806.
- ^ Джи Кью, Луо ZX, Юань С.Х., Wible JR, Чжан Дж. П., Георгий Дж. А. (апрель 2002 г.). «Самое раннее известное млекопитающее эвтерианской породы». Природа. 416 (6883): 816–22. Bibcode:2002Натура.416..816J. Дои:10.1038 / 416816a. PMID 11976675. S2CID 4330626.
- ^ Новачек MJ, Rougier GW, Wible JR, McKenna MC, Dashzeveg D, Horovitz I (октябрь 1997 г.). «Надлобковые кости у здоровых млекопитающих из позднего мела Монголии». Природа. 389 (6650): 483–6. Bibcode:1997Натура.389..483Н. Дои:10.1038/39020. PMID 9333234. S2CID 205026882.
- ^ Power ML, Schulkin J (2012). «Эволюция живорождения у млекопитающих». Эволюция плаценты человека. Балтимор: Издательство Университета Джона Хопкинса. п. 68. ISBN 978-1-4214-0643-5.
- ^ Rowe T, Rich TH, Vickers-Rich P, Springer M, Woodburne MO (январь 2008 г.). «Самый старый утконос и его влияние на время расхождения клад утконоса и ехидны». Труды Национальной академии наук Соединенных Штатов Америки. 105 (4): 1238–42. Bibcode:2008ПНАС..105.1238Р. Дои:10.1073 / pnas.0706385105. ЧВК 2234122. PMID 18216270.
- ^ Грант Т (1995). "Репродукция". Утконос: уникальное млекопитающее. Сидней: Университет Нового Южного Уэльса. п. 55. ISBN 978-0-86840-143-0. OCLC 33842474.
- ^ Goldman AS (июнь 2012 г.). «Эволюция иммунных функций молочной железы и защита младенца». Медицина грудного вскармливания. 7 (3): 132–42. Дои:10.1089 / bfm.2012.0025. PMID 22577734.
- ^ а б Роза К.Д. (2006). Начало эры млекопитающих. Балтимор: Издательство Университета Джона Хопкинса. С. 82–83. ISBN 978-0-8018-8472-6. OCLC 646769601.
- ^ Бринк А.С. (1955). "Исследование скелета Диадемодон". Palaeontologia Africana. 3: 3–39.
- ^ Кемп Т.С. (1982). Млекопитающие рептилии и происхождение млекопитающих. Лондон: Academic Press. п. 363. ISBN 978-0-12-404120-2. OCLC 8613180.
- ^ Эстес Р. (1961). «Анатомия черепа цинодонтовой рептилии. Thrinaxodon liorhinus". Вестник Музея сравнительной зоологии (1253): 165–180.
- ^ "Тринаксодон: Новое млекопитающее ". National Geographic Daily News. 11 февраля 2009 г.. Получено 26 августа, 2012.
- ^ а б Байдек П., Кварнстрём М., Овоцкий К., Сулей Т., Сенников А.Г., Голубев В.К., Недзведский Г. (2015). «Микробиота и пищевые остатки, включая возможные свидетельства наличия волос до млекопитающих в копролитах верхней перми из России». Lethaia. 49 (4): 455–477. Дои:10.1111 / лет.12156.
- ^ Бота-Бринк Дж, Ангельчик К.Д. (2010). «Объясняют ли необычайно высокие темпы роста пермо-триасовых дицинодонтов (Therapsida, Anomodontia) их успех до и после вымирания в конце пермского периода?». Зоологический журнал Линнеевского общества. 160 (2): 341–365. Дои:10.1111 / j.1096-3642.2009.00601.x.
- ^ Пол Г.С. (1988). Хищные динозавры мира. Нью-Йорк: Саймон и Шустер. п.464. ISBN 978-0-671-61946-6. OCLC 18350868.
- ^ Уотсон Дж. М., Грейвс Дж. А. (1988). «Монотремные клеточные циклы и эволюция гомеотермии». Австралийский зоологический журнал. 36 (5): 573–584. Дои:10.1071 / ZO9880573.
- ^ Макнаб Б.К. (1980). «Энергетика и пределы умеренного распространения броненосцев». Журнал маммологии. 61 (4): 606–627. Дои:10.2307/1380307. JSTOR 1380307.
- ^ Kielan-Jaworowska Z, Hurum JH (2006). «Положение конечностей у ранних млекопитающих: раскидистое или парасагиттальное» (PDF). Acta Palaeontologica Polonica. 51 (3): 10237–10239.
- ^ Lillegraven JA, Kielan-Jaworowska Z, Clemens WA (1979). Мезозойские млекопитающие: первые две трети истории млекопитающих. Калифорнийский университет Press. п. 321. ISBN 978-0-520-03951-3. OCLC 5910695.
- ^ Офтедал ОТ (июль 2002 г.). «Молочная железа и ее происхождение в процессе эволюции синапсидов». Журнал биологии и неоплазии молочных желез. 7 (3): 225–52. Дои:10.1023 / А: 1022896515287. PMID 12751889. S2CID 25806501.
- ^ Офтедал ОТ (июль 2002 г.). «Происхождение лактации как источника воды для яиц с пергаментной скорлупой». Журнал биологии и неоплазии молочных желез. 7 (3): 253–66. Дои:10.1023 / А: 1022848632125. PMID 12751890. S2CID 8319185.
- ^ а б Сахни С., Бентон М.Дж., Ферри, штат Пенсильвания (август 2010 г.). «Связи между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше». Письма о биологии. 6 (4): 544–7. Дои:10.1098 / рсбл.2009.1024. ЧВК 2936204. PMID 20106856.
- ^ Смит Ф.А., Бойер А.Г., Браун Дж. Х., Коста Д. П., Даян Т., Эрнест С. К. и др. (Ноябрь 2010 г.). «Эволюция максимальных размеров тела наземных млекопитающих». Наука. 330 (6008): 1216–9. Bibcode:2010Научный ... 330.1216S. CiteSeerX 10.1.1.383.8581. Дои:10.1126 / science.1194830. PMID 21109666. S2CID 17272200.
- ^ Симмонс Н.Б., Сеймур К.Л., Хаберсетцер Дж., Ганнелл Г.Ф. (февраль 2008 г.). «Примитивная летучая мышь раннего эоцена из Вайоминга и эволюция полета и эхолокации». Природа. 451 (7180): 818–21. Bibcode:2008Натура.451..818S. Дои:10.1038 / природа06549. HDL:2027.42/62816. PMID 18270539. S2CID 4356708.
- ^ Бининда-Эмондс OR, Cardillo M, Jones KE, MacPhee RD, Beck RM, Grenyer R и др. (Март 2007 г.). «Отсроченный рост современных млекопитающих» (PDF). Природа. 446 (7135): 507–12. Bibcode:2007Натура.446..507Б. Дои:10.1038 / природа05634. PMID 17392779. S2CID 4314965.
- ^ а б Wible JR, Rougier GW, Novacek MJ, Asher RJ (июнь 2007 г.). «Меловые эвтерианы и лавразийское происхождение плацентарных млекопитающих вблизи границы К / Т». Природа. 447 (7147): 1003–6. Bibcode:2007 Натур.447.1003W. Дои:10.1038 / природа05854. PMID 17581585. S2CID 4334424.
- ^ О'Лири М.А., Блох Дж. И., Флинн Дж. Дж., Годен Т. Дж., Джалломбардо А., Джаннини Н. П. и др. (Февраль 2013). "Предок плацентарных млекопитающих и пост-K-Pg-излучение плаценты". Наука. 339 (6120): 662–7. Bibcode:2013Наука ... 339..662O. Дои:10.1126 / science.1229237. PMID 23393258. S2CID 206544776.
- ^ Халлидей Т.Дж., Апчерч П., Госвами А. (февраль 2017 г.). «Разрешение родства плацентарных млекопитающих палеоцена». Биологические обзоры Кембриджского философского общества. 92 (1): 521–550. Дои:10.1111 / brv.12242. ЧВК 6849585. PMID 28075073.
- ^ Халлидей Т.Дж., Апчерч П., Госвами А. (июнь 2016 г.). «Евтерийцы испытали повышенные темпы эволюции сразу после массового вымирания в мелово-палеогеновый период». Ход работы. Биологические науки. 283 (1833): 20153026. Дои:10.1098 / rspb.2015.3026. ЧВК 4936024. PMID 27358361.
- ^ а б c Ни Х, Гебо Д.Л., Дагосто М., Мэн Дж., Таффоро П., Флинн Дж. Дж., Борода К.С. (июнь 2013 г.). «Самый старый известный скелет приматов и ранняя эволюция гаплорин». Природа. 498 (7452): 60–4. Bibcode:2013Натура 498 ... 60N. Дои:10.1038 / природа12200. PMID 23739424. S2CID 4321956.
- ^ Ромер С.А., Парсонс Т.С. (1977). Тело позвоночного. Филадельфия: Холт-Сондерс Интернэшнл. С. 129–145. ISBN 978-0-03-910284-5. OCLC 60007175.
- ^ Purves WK, Sadava DE, Orians GH, Helle HC (2001). Жизнь: Наука биологии (6 изд.). Нью-Йорк: Sinauer Associates, Inc., стр. 593. ISBN 978-0-7167-3873-2. OCLC 874883911.
- ^ Антвал Н., Джоши Л., Такер А.С. (январь 2013 г.). «Эволюция среднего уха и челюсти млекопитающих: адаптации и новые структуры». Журнал анатомии. 222 (1): 147–60. Дои:10.1111 / j.1469-7580.2012.01526.x. ЧВК 3552421. PMID 22686855.
- ^ ван Нивельт А.Ф., Смит К.К. (2005). «Заменять или не заменять: значение снижения функциональной замены зубов у сумчатых и плацентарных млекопитающих». Палеобиология. 31 (2): 324–346. Дои:10.1666 / 0094-8373 (2005) 031 [0324: trontr] 2.0.co; 2.
- ^ Libertini, G .; Феррара, Н. (2016). «Старение многолетних клеток и частей органов согласно парадигме запрограммированного старения». ВОЗРАСТ. 38 (35): 35. Дои:10.1007 / s11357-016-9895-0. ЧВК 5005898. PMID 26957493.
- ^ Мао Ф, Ван И, Мэн Дж (2015). «Систематическое исследование микроструктур зубной эмали Lambdopsalis bulla (Multituberculate, Mammalia) - значение для биологии и филогении мультитуберкулеза». PLOS ONE. 10 (5): e0128243. Bibcode:2015PLoSO..1028243M. Дои:10.1371 / journal.pone.0128243. ЧВК 4447277. PMID 26020958.
- ^ Осборн Х.Ф. (1900). "Происхождение млекопитающих, III. Затылочные мыщелки рептильного трехраздельного типа". Американский натуралист. 34 (408): 943–947. Дои:10.1086/277821. JSTOR 2453526.
- ^ Кромптон А.В., Дженкинс-младший Ф.А. (1973). «Млекопитающие от рептилий: обзор происхождения млекопитающих». Ежегодный обзор наук о Земле и планетах. 1: 131–155. Bibcode:1973AREPS ... 1..131C. Дои:10.1146 / annurev.ea.01.050173.001023.
- ^ а б Power ML, Schulkin J (2013). Эволюция плаценты человека. Балтимор: Издательство Университета Джона Хопкинса. С. 1890–1891. ISBN 978-1-4214-0643-5. OCLC 940749490.
- ^ Линденфорс, Патрик; Гиттлман, Джон Л .; Джонс, Кейт (2007), «Половой размерный диморфизм у млекопитающих», Пол, размер и гендерные роли, стр., с. 16–26, ISBN 978-0-19-920878-4, получено 2020-10-08
- ^ Dierauf LA, Gulland FM (2001). Справочник CRC по медицине морских млекопитающих: здоровье, болезни и реабилитация (2-е изд.). Бока-Ратон: CRC Press. п. 154. ISBN 978-1-4200-4163-7. OCLC 166505919.
- ^ Луи Дж. Х., Хансен Д. В., Кригштейн А. Р. (июль 2011 г.). «Развитие и эволюция неокортекса человека». Клетка. 146 (1): 18–36. Дои:10.1016 / j.cell.2011.06.030. ЧВК 3610574. PMID 21729779.
- ^ Киллер CE (июнь 1933 г.). "Отсутствие мозолистого тела как менделизирующего характера у домашней мыши". Труды Национальной академии наук Соединенных Штатов Америки. 19 (6): 609–11. Bibcode:1933ПНАС ... 19..609К. Дои:10.1073 / пнас.19.6.609. JSTOR 86284. ЧВК 1086100. PMID 16587795.
- ^ Левицкий М.Г. (2013). «Механика дыхания». Легочная физиология (8-е изд.). Нью-Йорк: McGraw-Hill Medical. ISBN 978-0-07-179313-1. OCLC 940633137.
- ^ а б Умеш КБ (2011). «Легочная анатомия и физиология». Справочник по механической вентиляции (1-е изд.). Нью-Дели: издательство Jaypee Brothers Medical Publishing. п. 12. ISBN 978-93-80704-74-6. OCLC 945076700.
- ^ Standring S, Borley NR (2008). Анатомия Грея: анатомические основы клинической практики (40 изд.). Лондон: Черчилль Ливингстон. С. 960–962. ISBN 978-0-8089-2371-8. OCLC 213447727.
- ^ Беттс Дж. Г., Десе П., Джонсон Э, Джонсон Дж. Э., Корол О, Круз Д., По Б., Мудрый Дж. А., Уомбл М., Янг К. А. (2013). Анатомия и физиология. Хьюстон: Издательство Университета Райса. С. 787–846. ISBN 978-1-938168-13-0. OCLC 898069394.
- ^ а б c d е ж грамм час я Feldhamer GA, Drickamer LC, Vessey SH, Merritt JH, Krajewski C (2007). Маммология: адаптация, разнообразие, экология (3-е изд.). Балтимор: Издательство Университета Джона Хопкинса. ISBN 978-0-8018-8695-9. OCLC 124031907.
- ^ Тинкер С.В. (1988). Киты мира. Brill Archive. п. 51. ISBN 978-0-935848-47-2.
- ^ Ромер А.С. (1959). История позвоночных (4-е изд.). Чикаго: Издательство Чикагского университета. ISBN 978-0-226-72490-4.
- ^ де Мюисон С., Ланге-Бадре Б. (1997). «Плотоядные дентальные приспособления у трибосфенических млекопитающих и филогенетическая реконструкция». Lethaia. 30 (4): 353–366. Дои:10.1111 / j.1502-3931.1997.tb00481.x.
- ^ а б Лангер П. (июль 1984 г.). «Сравнительная анатомия желудка у травоядных млекопитающих». Ежеквартальный журнал экспериментальной физиологии. 69 (3): 615–25. Дои:10.1113 / expphysiol.1984.sp002848. PMID 6473699.
- ^ Воан Т.А., Райан Дж. М., Чаплевски Н.Дж. (2011). «Периссодактиля». Маммология (5-е изд.). Джонс и Бартлетт. п. 322. ISBN 978-0-7637-6299-5. OCLC 437300511.
- ^ Цветок WH, Lydekker R (1946). Введение в изучение живых и вымерших млекопитающих. Лондон: Адам и Чарльз Блэк. п. 496. ISBN 978-1-110-76857-8.
- ^ Sreekumar S (2010). Базовая физиология. PHI Learning Pvt. Ltd. С. 180–181. ISBN 978-81-203-4107-4.
- ^ Cheifetz AS (2010). Оксфордский американский справочник по гастроэнтерологии и гепатологии. Оксфорд: Издательство Оксфордского университета, США. п. 165. ISBN 978-0-19-983012-1.
- ^ Кунц Э (2008). Гепатология: Учебник и атлас. Германия: Springer. п. 38. ISBN 978-3-540-76838-8.
- ^ Ортис Р.М. (июнь 2001 г.). «Осморегуляция у морских млекопитающих». Журнал экспериментальной биологии. 204 (Pt 11): 1831–44. PMID 11441026.
- ^ а б c Роман А.С., Парсонс Т.С. (1977). Тело позвоночного. Филадельфия: Холт-Сондерс Интернэшнл. С. 396–399. ISBN 978-0-03-910284-5.
- ^ Биологические обзоры - Cambridge Journals
- ^ Докинз Р., Вонг И (2016). Сказка предков: паломничество к заре эволюции (2-е изд.). Бостон: Mariner Books. п. 281. ISBN 978-0-544-85993-7.
- ^ а б c Fitch WT (2006 г.). «Производство вокализаций у млекопитающих» (PDF). В Браун К (ред.). Энциклопедия языка и лингвистики. Оксфорд: Эльзевир. С. 115–121.
- ^ Ланжевен П., Барклай Р.М. (1990). "Hypsignathus monstrosus". Виды млекопитающих. 357 (357): 1–4. Дои:10.2307/3504110. JSTOR 3504110.
- ^ Weissengruber GE, Forstenpointner G, Peters G, Kübber-Heiss A, Fitch WT (сентябрь 2002 г.). «Подъязычный аппарат и глотка у льва (Panthera leo), ягуара (Panthera onca), тигра (Panthera tigris), гепарда (Acinonyxjubatus) и домашней кошки (Felis silvestris f. Catus)». Журнал анатомии. 201 (3): 195–209. Дои:10.1046 / j.1469-7580.2002.00088.x. ЧВК 1570911. PMID 12363272.
- ^ Stoeger AS, Heilmann G, Zeppelzauer M, Ganswindt A, Hensman S, Charlton BD (2012). «Визуализация звуковой эмиссии вокализации слона: свидетельство двух типов грохота». PLOS ONE. 7 (11): e48907. Bibcode:2012PLoSO ... 748907S. Дои:10.1371 / journal.pone.0048907. ЧВК 3498347. PMID 23155427.
- ^ Кларк CW (2004). «Инфразвуковые звуки усатого кита: естественная изменчивость и функция». Журнал Акустического общества Америки. 115 (5): 2554. Bibcode:2004ASAJ..115.2554C. Дои:10.1121/1.4783845.
- ^ а б Доусон Т.Дж., Вебстер К.Н., Мэлони СК (февраль 2014 г.). «Мех млекопитающих в открытой среде; обязательно ли конфликтуют крипсис и тепловые потребности? Сравнение белого медведя и сумчатой коалы». Журнал сравнительной физиологии B. 184 (2): 273–84. Дои:10.1007 / s00360-013-0794-8. PMID 24366474. S2CID 9481486.
- ^ Сломинский А., Тобин Д. Д., Шибахара С., Вортсман Дж. (Октябрь 2004 г.). «Пигментация меланина в коже млекопитающих и ее гормональная регуляция». Физиологические обзоры. 84 (4): 1155–228. Дои:10.1152 / физрев.00044.2003. PMID 15383650. S2CID 21168932.
- ^ Хилтон младший Б. (1996). "Дикая природа Южной Каролины". Цвета Животных. Хилтон Понд Центр. 43 (4): 10–15. Получено 26 ноября 2011.
- ^ а б Прум Р.О., Торрес Р.Х. (май 2004 г.). «Структурная окраска кожи млекопитающих: конвергентная эволюция когерентно рассеивающих массивов кожного коллагена» (PDF). Журнал экспериментальной биологии. 207 (Пт 12): 2157–72. Дои:10.1242 / jeb.00989. HDL:1808/1599. PMID 15143148. S2CID 8268610.
- ^ Суутари М., Маянева М., Фьюер Д.П., Вурин Б., Айелло А., Фридл Т. и др. (Март 2010 г.). «Молекулярные доказательства разнообразного сообщества зеленых водорослей, растущих в волосах ленивцев, и специфической ассоциации с Trichophilus welckeri (Chlorophyta, Ulvophyceae)». BMC Эволюционная биология. 10 (86): 86. Дои:10.1186/1471-2148-10-86. ЧВК 2858742. PMID 20353556.
- ^ Каро Т. (2005). «Адаптивное значение окраски млекопитающих». Бионаука. 55 (2): 125–136. Дои:10.1641 / 0006-3568 (2005) 055 [0125: tasoci] 2.0.co; 2.
- ^ Миллс Л.С., Зимова М., Ойлер Дж., Бегущий С., Абацоглу Дж. Т., Лукач П.М. (апрель 2013 г.). «Несоответствие камуфляжа сезонной окраске шерсти из-за уменьшения продолжительности снежного покрова». Труды Национальной академии наук Соединенных Штатов Америки. 110 (18): 7360–5. Bibcode:2013ПНАС..110.7360М. Дои:10.1073 / pnas.1222724110. ЧВК 3645584. PMID 23589881.
- ^ Каро Т. (февраль 2009 г.). «Контрастная окраска наземных млекопитающих». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 364 (1516): 537–48. Дои:10.1098 / rstb.2008.0221. ЧВК 2674080. PMID 18990666.
- ^ Плавкан Ю.М. (2001). «Половой диморфизм в эволюции приматов». Американский журнал физической антропологии. Дополнение 33 (33): 25–53. Дои:10.1002 / ajpa.10011. PMID 11786990.
- ^ Брэдли Б.Дж., Джеральд М.С., Виддиг А., Манди Н.И. (2012). «Вариация цвета шерсти и экспрессия генов пигментации у макак-резусов (Macaca Mulatta)» (PDF). Журнал эволюции млекопитающих. 20 (3): 263–270. Дои:10.1007 / s10914-012-9212-3. S2CID 13916535. Архивировано из оригинал (PDF) на 2015-09-24.
- ^ Каро Т, Иззо А., Райнер Р.К., Уокер Х., Станкович Т. (апрель 2014 г.). «Функция полосок зебры». Nature Communications. 5: 3535. Bibcode:2014 НатКо ... 5,3535C. Дои:10.1038 / ncomms4535. PMID 24691390. S2CID 9849814.
- ^ Ломбарди Дж. (30 ноября 1998 г.). Сравнительное воспроизводство позвоночных. Springer Science & Business Media. ISBN 978-0-7923-8336-9.
- ^ Tyndale-Biscoe H, Renfree M (30 января 1987 г.). Репродуктивная физиология сумчатых. Издательство Кембриджского университета. ISBN 978-0-521-33792-2.
- ^ Джонстон С.Д., Смит Б., Пайн М., Стензель Д., Холт В.В. (2007). «Односторонняя эякуляция пучками спермы ехидны» (PDF). Американский натуралист. 170 (6): E162 – E164. Дои:10.1086/522847. PMID 18171162.
- ^ Максвелл К.Э. (2013). Сексуальный императив: эволюционная история сексуального выживания. Springer. С. 112–113. ISBN 978-1-4899-5988-1.
- ^ Воан Т.А., Райан Дж. П., Чаплевски Н.Дж. (2011). Маммология. Издательство "Джонс и Бартлетт". п. 387. ISBN 978-0-03-025034-7.
- ^ а б Хоффман Э.А., Роу ТБ (сентябрь 2018 г.). «Перинаты юрских стволовых млекопитающих и происхождение размножения и роста млекопитающих». Природа. 561 (7721): 104–108. Bibcode:2018Натура.561..104H. Дои:10.1038 / s41586-018-0441-3. PMID 30158701. S2CID 205570021.
- ^ Уоллис М.К., Уотерс П.Д., Дельбридж М.Л., Кирби П.Дж., Паск А.Дж., Грюцнер Ф. и др. (2007). «Определение пола у утконоса и ехидны: аутосомное расположение SOX3 подтверждает отсутствие SRY у монотрем». Хромосомные исследования. 15 (8): 949–59. Дои:10.1007 / s10577-007-1185-3. PMID 18185981. S2CID 812974.
- ^ Маршалл Грейвс Дж. А. (2008). «Странные геномы животных и эволюция половых и половых хромосом позвоночных» (PDF). Ежегодный обзор генетики. 42: 565–86. Дои:10.1146 / annurev.genet.42.110807.091714. PMID 18983263. Архивировано из оригинал (PDF) на 2012-09-04.
- ^ Новачек MJ, Rougier GW, Wible JR, McKenna MC, Dashzeveg D, Horovitz I (октябрь 1997 г.). «Надлобковые кости у здоровых млекопитающих из позднего мела Монголии». Природа. 389 (6650): 483–6. Bibcode:1997Натура.389..483Н. Дои:10.1038/39020. PMID 9333234. S2CID 205026882.
- ^ Морган С (2005). «Поведение и образ жизни млекопитающих». Млекопитающие. Чикаго: Рейнтри. п. 6. ISBN 978-1-4109-1050-9. OCLC 53476660.
- ^ Верма П.С., Панди Б.П. (2013). Книга I по биологии ISC для класса XI. Нью-Дели: С. Чанд и компания. п. 288. ISBN 978-81-219-2557-0.
- ^ Офтедал ОТ (июль 2002 г.). «Молочная железа и ее происхождение в процессе эволюции синапсидов». Журнал биологии и неоплазии молочных желез. 7 (3): 225–52. Дои:10.1023 / а: 1022896515287. PMID 12751889. S2CID 25806501.
- ^ Крокенбергер А (2006). "Кормление грудью". В Dickman CR, Armati PJ, Hume ID (ред.). Сумчатые. п. 109. ISBN 9781139457422.
- ^ Шулькин Дж, Power ML (2016). Молоко: биология лактации. Издательство Университета Джона Хопкинса. п. 66. ISBN 9781421420424.
- ^ Томпсон К.В., Бейкер AJ, Бейкер AM (2010). «Отцовский уход и развитие поведения у млекопитающих в неволе». В Baer CK, Kleiman DG, Thompson KV (ред.). Дикие млекопитающие в неволе Принципы и методы управления зоопарком (2-е изд.). Издательство Чикагского университета. п. 374. ISBN 9780226440118.
- ^ Кэмпбелл Н. А., Рис Дж. Б. (2002). Биология (6 изд.). Бенджамин Каммингс. п.845. ISBN 978-0-8053-6624-2. OCLC 47521441.
- ^ Buffenstein R, Yahav S (1991). "Голый землекоп Герероцефал глабер эндотермическое, но пойкилотермное млекопитающее? ". Журнал термобиологии. 16 (4): 227–232. Дои:10.1016/0306-4565(91)90030-6.
- ^ Шмидт-Нильсен К., Дюк Дж. Б. (1997). «Температурные эффекты». Физиология животных: адаптация и окружающая среда (5-е изд.). Кембридж. п. 218. ISBN 978-0-521-57098-5. OCLC 35744403.
- ^ а б Лоренцини А., Джонсон Ф. Б., Оливер А., Трезини М., Смит Дж. С., Хдеиб М. и др. (2009). «Значительная корреляция долголетия видов с распознаванием двухцепочечных разрывов ДНК, но не с длиной теломер». Механизмы старения и развития. 130 (11–12): 784–92. Дои:10.1016 / j.mad.2009.10.004. ЧВК 2799038. PMID 19896964.
- ^ Харт Р. У., Сетлоу РБ (июнь 1974 г.). «Корреляция между эксцизионным восстановлением дезоксирибонуклеиновой кислоты и продолжительностью жизни у ряда видов млекопитающих». Труды Национальной академии наук Соединенных Штатов Америки. 71 (6): 2169–73. Bibcode:1974PNAS ... 71.2169H. Дои:10.1073 / pnas.71.6.2169. ЧВК 388412. PMID 4526202.
- ^ Ма С., Упнея А., Галецки А., Цай Ю. М., Бурант С. Ф., Раскинд С. и др. (Ноябрь 2016 г.). «Профилирование на основе клеточных культур у млекопитающих показывает, что восстановление ДНК и метаболизм являются определяющими факторами долголетия видов». eLife. 5. Дои:10.7554 / eLife.19130. ЧВК 5148604. PMID 27874830.
- ^ Grube K, Bürkle A (декабрь 1992 г.). «Активность поли (АДФ-рибозы) полимеразы в мононуклеарных лейкоцитах 13 видов млекопитающих коррелирует с видоспецифичной продолжительностью жизни». Труды Национальной академии наук Соединенных Штатов Америки. 89 (24): 11759–63. Bibcode:1992PNAS ... 8911759G. Дои:10.1073 / пнас.89.24.11759. ЧВК 50636. PMID 1465394.
- ^ Фрэнсис А.А., Ли У.Х., Реган ДжейДи (июнь 1981 г.) «Связь эксцизионной репарации ДНК ультрафиолетовых повреждений с максимальной продолжительностью жизни млекопитающих». Механизмы старения и развития. 16 (2): 181–9. Дои:10.1016/0047-6374(81)90094-4. PMID 7266079. S2CID 19830165.
- ^ Третон Дж. А., Куртуа И. (март 1982 г.). «Корреляция между эксцизионной репарацией ДНК и продолжительностью жизни млекопитающих в эпителиальных клетках хрусталика». Международные отчеты по клеточной биологии. 6 (3): 253–60. Дои:10.1016/0309-1651(82)90077-7. PMID 7060140.
- ^ Маслански CJ, Williams GM (февраль 1985 г.). «Ультрафиолетовый свет индуцировал репарационный синтез ДНК в гепатоцитах видов разной продолжительности жизни». Механизмы старения и развития. 29 (2): 191–203. Дои:10.1016/0047-6374(85)90018-1. PMID 3974310. S2CID 23988416.
- ^ «Нога и ступни». Адаптации птичьего скетета. Архивировано из оригинал на 2008-04-04. Получено 3 августа 2008.
- ^ Уокер ВФ, Homberger DG (1998). Анатомия и расслоение плода свиньи (5-е изд.). Нью-Йорк: В. Х. Фриман и компания. п. 3. ISBN 978-0-7167-2637-1. OCLC 40576267.
- ^ Орр CM (ноябрь 2005 г.). «Муравьед, ходящий на костяшках пальцев: тест на адаптацию к предполагаемым особенностям ходьбы на костяшках пальцев африканских гоминид». Американский журнал физической антропологии. 128 (3): 639–58. Дои:10.1002 / ajpa.20192. PMID 15861420.
- ^ Fish FE, Frappell PB, Baudinette RV, MacFarlane PM (февраль 2001 г.). «Энергетика наземного передвижения утконоса Ornithorhynchus anatinus» (PDF). Журнал экспериментальной биологии. 204 (Pt 4): 797–803. PMID 11171362.
- ^ Дхингра П. (2004). "Сравнительное двуногие - как остальная часть животного мира ходит на двух ногах". Антропологическая наука. 131 (231).
- ^ Александр Р.М. (май 2004 г.). «Двуногие животные и их отличия от человека». Журнал анатомии. 204 (5): 321–30. Дои:10.1111 / j.0021-8782.2004.00289.x. ЧВК 1571302. PMID 15198697.
- ^ а б Dagg AI (1973). «Походки у млекопитающих». Обзор млекопитающих. 3 (4): 135–154. Дои:10.1111 / j.1365-2907.1973.tb00179.x.
- ^ Робертс Т.Д. (1995). Понимание баланса: механика позы и передвижения. Сан-Диего: Нельсон Торнс. п. 211. ISBN 978-1-56593-416-0. OCLC 33167785.
- ^ а б c Картмилл М (1985). "Альпинизм". В Hildebrand M, Bramble DM, Liem KF, Wake DB (ред.). Функциональная морфология позвоночных. Кембридж: Белкнап Пресс. С. 73–88. ISBN 978-0-674-32775-7. OCLC 11114191.
- ^ Верн К. (2001). «Планерное исполнение северной летяги (Glaucomys sabrinus) в зрелых смешанных лесах Восточной Канады » (PDF). Журнал маммологии. 82 (4): 1026–1033. Дои:10.1644 / 1545-1542 (2001) 082 <1026: GPOTNF> 2.0.CO; 2.
- ^ Барба Л.А. (октябрь 2011 г.). «Летучие мыши - единственные летающие млекопитающие». Био-воздушное движение. Получено 20 мая 2016.
- ^ «Летучие мыши в полете показывают неожиданную аэродинамику». ScienceDaily. 2007. Получено 12 июля, 2016.
- ^ Hedenström A, Johansson LC (март 2015 г.). «Полет летучей мыши: аэродинамика, кинематика и морфология полета» (PDF). Журнал экспериментальной биологии. 218 (Pt 5): 653–63. Дои:10.1242 / jeb.031203. PMID 25740899. S2CID 21295393.
- ^ «Летучие мыши экономят энергию, втягивая крылья при движении вверх». ScienceDaily. 2012. Получено 12 июля, 2016.
- ^ Тащек К (2008). Тусовка с летучими мышами: эко-летучие мыши, вампиры и кинозвезды. Альбукерке: Издательство Университета Нью-Мексико. п. 14. ISBN 978-0-8263-4403-8. OCLC 191258477.
- ^ Стербинг-Д'Анджело С., Чадха М., Чиу С., Фальк Б., Сиань В., Барсело Дж. И др. (Июль 2011 г.). «Датчики крыла летучей мыши поддерживают управление полетом». Труды Национальной академии наук Соединенных Штатов Америки. 108 (27): 11291–6. Bibcode:2011ПНАС..10811291С. Дои:10.1073 / pnas.1018740108. ЧВК 3131348. PMID 21690408.
- ^ Дамиани, Р., 2003, Самые ранние свидетельства роения цинодонтов, Издательство Королевского общества, Том 270, Выпуск 1525
- ^ Шимер HW (1903). «Адаптация к водным, древесным, фоссориальным и поверхностным привычкам млекопитающих. III. Фоссориальные адаптации». Американский натуралист. 37 (444): 819–825. Дои:10.1086/278368. JSTOR 2455381.
- ^ Стэнхоуп, М. Дж .; Waddell, V.G .; и другие. (1998). «Молекулярные доказательства множественного происхождения насекомоядных и нового отряда эндемичных африканских насекомоядных млекопитающих». Труды Национальной академии наук. 95 (17): 9967–9972. Bibcode:1998PNAS ... 95.9967S. Дои:10.1073 / пнас.95.17.9967. ЧВК 21445. PMID 9707584.
- ^ Перри Д.А. (1949). «Анатомические основы плавания у китов». Журнал зоологии. 119 (1): 49–60. Дои:10.1111 / j.1096-3642.1949.tb00866.x.
- ^ Fish FE, Хуэй, Калифорния (1991). «Плавание с дельфинами - обзор» (PDF). Обзор млекопитающих. 21 (4): 181–195. Дои:10.1111 / j.1365-2907.1991.tb00292.x. Архивировано из оригинал (PDF) 29 августа 2006 г.
- ^ Марш Х (1989). "Глава 57: Дюгоньи" (PDF). Фауна Австралии. 1. Канберра: публикации правительства Австралии. ISBN 978-0-644-06056-1. OCLC 27492815. Архивировано 11 мая 2013 года.CS1 maint: BOT: статус исходного URL-адреса неизвестен (связь)
- ^ а б Берта А. (апрель 2012 г.). «Разнообразие ластоногих: эволюция и адаптации». Возвращение в море: жизнь и время эволюции морских млекопитающих. Калифорнийский университет Press. С. 62–64. ISBN 978-0-520-27057-2.
- ^ а б Fish FE, Hurley J, Costa DP (февраль 2003 г.). «Маневренность морского льва Zalophus californianus: поворотные характеристики нестабильной конструкции тела». Журнал экспериментальной биологии. 206 (Pt 4): 667–74. Дои:10.1242 / jeb.00144. PMID 12517984.
- ^ а б Ридман М (1990). Ластоногие: тюлени, морские львы и моржи. Калифорнийский университет Press. ISBN 978-0-520-06497-3. OCLC 19511610.
- ^ Рыбная ИП (1996). «Переход от тяги к подъемной силе в плавании млекопитающих». Интегративная и сравнительная биология. 36 (6): 628–641. Дои:10.1093 / icb / 36.6.628.
- ^ Рыбный ИП (2000). «Биомеханика и энергетика водных и полуводных млекопитающих: от утконоса до кита» (PDF). Физиологическая и биохимическая зоология. 73 (6): 683–98. CiteSeerX 10.1.1.734.1217. Дои:10.1086/318108. PMID 11121343. Архивировано из оригинал (PDF) на 2016-08-04.
- ^ Элтрингем СК (1999). "Анатомия и психология". Бегемоты. Лондон: T&AD Poyser Ltd., стр. 8. ISBN 978-0-85661-131-5. OCLC 42274422.
- ^ "Бегемот Бегемот амфибий". Национальная география. В архиве из оригинала 25.11.2014. Получено 30 апреля 2016.
- ^ а б Сейфарт Р.М., Чейни Д.Л., Марлер П. (1980). «Тревожные звонки обезьян Vervet: семантическое общение в свободном примате». Поведение животных. 28 (4): 1070–1094. Дои:10.1016 / S0003-3472 (80) 80097-2. S2CID 53165940.
- ^ Цубербюлер К (2001). "Тревожные сигналы хищников у обезьян Кэмпбелла, Cercopithecus campbelli". Поведенческая экология и социобиология. 50 (5): 414–442. Дои:10.1007 / s002650100383. JSTOR 4601985. S2CID 21374702.
- ^ Слаббекорн Х., Смит Т.Б. (апрель 2002 г.). «Пение птиц, экология и видообразование». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 357 (1420): 493–503. Дои:10.1098 / rstb.2001.1056. ЧВК 1692962. PMID 12028787.
- ^ Баннистер JL (2008). "Усатые киты (Mysticetes)". In F Perrin W, Würsig B, Thewissen JG (ред.). Энциклопедия морских млекопитающих (2-е изд.). Академическая пресса. С. 80–89. ISBN 978-0-12-373553-9.
- ^ Норрис С. (2002). «Существа культуры? В пользу культурных систем у китов и дельфинов» (PDF). Бионаука. 52 (1): 9–14. Дои:10.1641 / 0006-3568 (2002) 052 [0009: COCMTC] 2.0.CO; 2.
- ^ Boughman JW (февраль 1998 г.). «Обучение вокалу у больших остроносых летучих мышей». Ход работы. Биологические науки. 265 (1392): 227–33. Дои:10.1098 / rspb.1998.0286. ЧВК 1688873. PMID 9493408.
- ^ "Язык луговых собачек расшифрован учеными". CBC News. 21 июня 2013 г.. Получено 20 мая 2015.
- ^ Мэйелл Х (3 марта 2004 г.). «Слоны звонят в нерабочее время». Национальная география. Получено 15 ноября 2016.
- ^ Мейнард Смит Дж., Харпер Д. (2003). Сигналы животных. Оксфордская серия по экологии и эволюции. Издательство Оксфордского университета. С. 61–63. ISBN 978-0-19-852684-1. OCLC 54460090.
- ^ Компакт-диск Фитцгиббона, Fanshawe JH (1988). «Стоять в газелях Томсона: честный сигнал состояния» (PDF). Поведенческая экология и социобиология. 23 (2): 69–74. Дои:10.1007 / bf00299889. S2CID 2809268. Архивировано из оригинал (PDF) на 2014-02-25.
- ^ Бильдштейн К.Л. (май 1983 г.). "Почему белохвостые олени флагом хвосты". Американский натуралист. 121 (5): 709–715. Дои:10.1086/284096. JSTOR 2460873.
- ^ Гослинг Л.М. (январь 1982 г.). «Переоценка функции ароматической маркировки территорий». Zeitschrift für Tierpsychologie. 60 (2): 89–118. Дои:10.1111 / j.1439-0310.1982.tb00492.x.
- ^ Zala SM, Potts WK, Penn DJ (март 2004 г.). «Индикация запаха достоверно сигнализирует о здоровье и инфекции». Поведенческая экология. 15 (2): 338–44. Дои:10.1093 / beheco / arh022. HDL:10.1093 / beheco / arh022.
- ^ Джонсон Р.П. (август 1973 г.). «Ароматическая маркировка млекопитающих». Поведение животных. 21 (3): 521–535. Дои:10.1016 / S0003-3472 (73) 80012-0.
- ^ Шевилл В.Е., Макбрайд А.Ф. (1956). «Свидетельства эхолокации китообразных». Глубоководные исследования. 3 (2): 153–154. Bibcode:1956DSR ..... 3..153S. Дои:10.1016 / 0146-6313 (56) 90096-х.
- ^ Уилсон В., Мосс С. (2004). Томас Дж (ред.). Эхолокация у летучих мышей и дельфинов. Издательство Чикагского университета. п. 22. ISBN 978-0-226-79599-7. OCLC 50143737.
- ^ Au WW (1993). Сонар дельфинов. Springer-Verlag. ISBN 978-3-540-97835-0. OCLC 26158593.
- ^ Сандерс Дж. Г., Бейчман А. С., Роман Дж., Скотт Дж. Дж., Эмерсон Д., Маккарти Дж. Дж., Гиргис PR (сентябрь 2015 г.). «У усатых китов уникальный микробиом кишечника, сходный как с плотоядными, так и с травоядными». Nature Communications. 6: 8285. Bibcode:2015НатКо ... 6.8285S. Дои:10.1038 / ncomms9285. ЧВК 4595633. PMID 26393325.
- ^ Спиксман-младший (1996). «Энергетика и эволюция размеров тела у мелких наземных млекопитающих» (PDF). Симпозиумы Лондонского зоологического общества (69): 69–81.
- ^ а б Уилсон Д.Е., Берни Д., ред. (2001). Животное: подробное визуальное руководство по дикой природе мира (1-е изд.). DK Publishing. С. 86–89. ISBN 978-0-7894-7764-4. OCLC 46422124.
- ^ а б Ван Валкенбург Б (июль 2007 г.). «Дежавю: эволюция морфологии питания хищников». Интегративная и сравнительная биология. 47 (1): 147–63. Дои:10.1093 / icb / icm016. PMID 21672827.
- ^ Сакко Т., ван Валкенбург Б. (2004). «Экоморфологические показатели пищевого поведения медведей (Carnivora: Ursidae)». Журнал зоологии. 263 (1): 41–54. Дои:10.1017 / S0952836904004856.
- ^ Певица М.С., Бернейс Е.А. (2003). «Понимание всеядности требует поведенческой точки зрения». Экология. 84 (10): 2532–2537. Дои:10.1890/02-0397.
- ^ Хатсон Дж. М., Берк CC, Хейнс Дж. (2013-12-01). «Остеофагия и модификации костей у жирафа и других крупных копытных». Журнал археологической науки. 40 (12): 4139–4149. Дои:10.1016 / j.jas.2013.06.004.
- ^ "Почему кошки едят траву?". Pet MD. Получено 13 января 2017.
- ^ Гейзер Ф (2004). «Снижение скорости обмена веществ и температуры тела во время спячки и ежедневного оцепенения». Ежегодный обзор физиологии. 66: 239–74. Дои:10.1146 / annurev.physiol.66.032102.115105. PMID 14977403. S2CID 22397415.
- ^ Хамфрис М.М., Томас Д.В., Крамер Д.Л. (2003). «Роль доступности энергии в гибернации млекопитающих: подход с точки зрения затрат и выгод». Физиологическая и биохимическая зоология. 76 (2): 165–79. Дои:10.1086/367950. PMID 12794670. S2CID 14675451.
- ^ Барнс Б.М. (июнь 1989 г.). «Предотвращение замораживания у млекопитающих: температура тела ниже 0 градусов С в арктической спячке». Наука. 244 (4912): 1593–5. Bibcode:1989Научный ... 244.1593B. Дои:10.1126 / science.2740905. PMID 2740905.
- ^ Гейзер Ф (2010). «Праздник млекопитающих и птиц». В Navas CA, Carvalho JE (ред.). Праздник: молекулярные и физиологические аспекты. Прогресс в молекулярной и субклеточной биологии. 49. Springer-Verlag. С. 95–113. Дои:10.1007/978-3-642-02421-4. ISBN 978-3-642-02420-7.
- ^ Манн Дж., Паттерсон Э.М. (ноябрь 2013 г.). «Использование инструмента водными животными». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 368 (1630): 20120424. Дои:10.1098 / rstb.2012.0424. ЧВК 4027413. PMID 24101631.
- ^ Раффаэле П (2011). Среди обезьян: приключения по следам наших ближайших родственников. Нью-Йорк: Харпер. п. 83. ISBN 978-0-06-167184-5. OCLC 674694369.
- ^ Кёлер W (1925). Ментальность обезьян. Liveright. ISBN 978-0-87140-108-3. OCLC 2000769.
- ^ Макгоуэн Р.Т., Рен Т., Норлинг Й., Килинг Л.Дж. (май 2014 г.). «Позитивный эффект и обучение: изучение« эффекта Эврики »у собак». Познание животных. 17 (3): 577–87. Дои:10.1007 / с10071-013-0688-х. PMID 24096703. S2CID 15216926.
- ^ Карбовски Дж. (Май 2007 г.). «Глобальное и региональное метаболическое масштабирование мозга и его функциональные последствия». BMC Биология. 5 (18): 18. arXiv:0705.2913. Bibcode:2007arXiv0705.2913K. Дои:10.1186/1741-7007-5-18. ЧВК 1884139. PMID 17488526.
- ^ Марино Л. (июнь 2007 г.). "Мозги китообразных: насколько они водные?". Анатомическая запись. 290 (6): 694–700. Дои:10.1002 / ар.20530. PMID 17516433.
- ^ Галоп Г.Г. (январь 1970 г.). «Шимпанзе: самопознание». Наука. 167 (3914): 86–7. Bibcode:1970Sci ... 167 ... 86G. Дои:10.1126 / science.167.3914.86. PMID 4982211. S2CID 145295899.
- ^ Плотник Дж. М., де Ваал Ф. Б., Рейсс Д. (ноябрь 2006 г.). «Самопознание в азиатском слоне» (PDF). Труды Национальной академии наук Соединенных Штатов Америки. 103 (45): 17053–7. Bibcode:2006PNAS..10317053P. Дои:10.1073 / pnas.0608062103. ЧВК 1636577. PMID 17075063.
- ^ Роберт С (1986). «Онтогенез зеркального поведения у двух видов человекообразных обезьян». Американский журнал приматологии. 10 (2): 109–117. Дои:10.1002 / ajp.1350100202. PMID 31979488.
- ^ Валравен V, ван Эльсакер Л., Верхейен Р. (1995). «Реакция группы карликовых шимпанзе (Pan paniscus) на их зеркальные изображения: свидетельство самопознания». Приматы. 36: 145–150. Дои:10.1007 / bf02381922. S2CID 38985498.
- ^ Лики Р. (1994). «Происхождение разума». Происхождение человечества. Нью-Йорк: BasicBooks. п. 150. ISBN 978-0-465-05313-1. OCLC 30739453.
- ^ Арчер Дж (1992). Этология и человеческое развитие. Роуман и Литтлфилд. С. 215–218. ISBN 978-0-389-20996-6. OCLC 25874476.
- ^ а б Мартен К., Псаракос С. (1995). «Свидетельства самосознания афалины (Tursiops truncatus) ". В Parker ST, Mitchell R, Boccia M (ред.). Самосознание у животных и людей: перспективы развития. Кембридж: Издательство Кембриджского университета. С. 361–379. ISBN 978-0-521-44108-7. OCLC 28180680.
- ^ а б Дельфур Ф., Мартен К. (апрель 2001 г.). «Обработка зеркальных изображений трех видов морских млекопитающих: косаток (Orcinus orca), ложных косаток (Pseudorca crassidens) и калифорнийских морских львов (Zalophus californianus)». Поведенческие процессы. 53 (3): 181–190. Дои:10.1016 / s0376-6357 (01) 00134-6. PMID 11334706. S2CID 31124804.
- ^ Джарвис Дж. Ю (май 1981 г.). «Эусоциальность у млекопитающего: кооперативное разведение в колониях голых землекопов». Наука. 212 (4494): 571–3. Bibcode:1981Научный ... 212..571J. Дои:10.1126 / science.7209555. JSTOR 1686202. PMID 7209555. S2CID 880054.
- ^ Джейкобс Д.С. и др. (1991). "Структура колонии и иерархия доминирования землекопа Дамараленда, Cryptomys damarensis (Rodentia: Bathyergidae) из Намибии ". Журнал зоологии. 224 (4): 553–576. Дои:10.1111 / j.1469-7998.1991.tb03785.x.
- ^ Харди С.Б. (2009). Матери и другие люди: эволюционные истоки взаимопонимания. Бостон: Belknap Press издательства Гарвардского университета. С. 92–93.
- ^ Харлоу Х.Ф., Суоми С.Дж. (июль 1971 г.). «Социальное восстановление с помощью обезьян, выращенных в изоляции». Труды Национальной академии наук Соединенных Штатов Америки. 68 (7): 1534–8. Bibcode:1971ПНАС ... 68.1534Н. Дои:10.1073 / pnas.68.7.1534. ЧВК 389234. PMID 5283943.
- ^ ван Шайк CP (январь 1999 г.). «Социоэкология социальности деления-синтеза у орангутанов». Приматы; Журнал приматологии. 40 (1): 69–86. Дои:10.1007 / BF02557703. PMID 23179533. S2CID 13366732.
- ^ Арчи EA, Moss CJ, Alberts SC (март 2006 г.). «Узы, которые связывают: генетическое родство предсказывает разделение и слияние социальных групп у диких африканских слонов». Ход работы. Биологические науки. 273 (1586): 513–22. Дои:10.1098 / rspb.2005.3361. ЧВК 1560064. PMID 16537121.
- ^ Смит Дж. Э., Меменис СК, Холекамп К. Э. (2007). "Выбор партнера по рангу в обществе деления-слияния пятнистой гиены (Crocuta crocuta)" (PDF). Поведенческая экология и социобиология. 61 (5): 753–765. Дои:10.1007 / s00265-006-0305-y. S2CID 24927919. Архивировано из оригинал (PDF) на 2014-04-25.
- ^ Матоба Т., Куцукаке Н., Хасегава Т. (2013). Хейворд М (ред.). «Потирание и облизывание головы укрепляют социальные связи в группе африканских львов в неволе, Panthera leo». PLOS ONE. 8 (9): e73044. Bibcode:2013PLoSO ... 873044M. Дои:10.1371 / journal.pone.0073044. ЧВК 3762833. PMID 24023806.
- ^ Крутцен М., Барре Л.М., Коннор Р.К., Манн Дж., Шервин В.Б. (июль 2004 г.). "'О, отец, где ты? »- оценка отцовства в обществе диких афалин (Tursiops sp.) В заливе Шарк, Западная Австралия». Молекулярная экология. 13 (7): 1975–90. Дои:10.1111 / j.1365-294X.2004.02192.x. PMID 15189218. S2CID 4510393.
- ^ Мартин С. (1991). Тропические леса Западной Африки: экология - угрозы - сохранение (1-е изд.). Springer. Дои:10.1007/978-3-0348-7726-8. ISBN 978-3-0348-7726-8.
- ^ le Roux A, Cherry MI, Gygax L (5 мая 2009 г.). «Бдительность поведения и последствия приспособленности: сравнение одиночного кормления и обязательного группового кормления млекопитающих». Поведенческая экология и социобиология. 63 (8): 1097–1107. Дои:10.1007 / s00265-009-0762-1. S2CID 21961356.
- ^ Паладжи Э., Норшиа I (2015). Samonds KE (ред.). «Время мира: примирение среди деспотических видов (Lemur catta)». PLOS ONE. 10 (11): e0142150. Bibcode:2015PLoSO..1042150P. Дои:10.1371 / journal.pone.0142150. ЧВК 4646466. PMID 26569400.
- ^ Восток ML, Хофер H (2000). «Самцы пятнистых гиен (Crocuta crocuta) очередь за статусом в социальных группах, где преобладают женщины ». Поведенческая экология. 12 (15): 558–568. Дои:10.1093 / beheco / 12.5.558.
- ^ Сэмюэлс А., Силк Дж. Б., Родман П. (1984). «Изменения в ранге доминирования и репродуктивном поведении самцов шляпных макак (Макака излучает)". Поведение животных. 32 (4): 994–1003. Дои:10.1016 / с0003-3472 (84) 80212-2. S2CID 53186523.
- ^ Дельпьетро Х.А., Руссо Р.Г. (2002). "Наблюдения за обыкновенной летучей мышью-вампиром (Desmodus rotundus) и волосатая летучая мышь-вампир (Дифилла экаудата) в плену". Биология млекопитающих. 67 (2): 65–78. Дои:10.1078/1616-5047-00011.
- ^ Клейман Д.Г. (март 1977 г.). «Моногамия у млекопитающих». Ежеквартальный обзор биологии. 52 (1): 39–69. Дои:10.1086/409721. PMID 857268. S2CID 25675086.
- ^ Холланд Б., Райс В. Р. (февраль 1998 г.). "Перспектива: сексуальный отбор изгнания: антагонистическое соблазнение против сопротивления" (PDF). Эволюция; Международный журнал органической эволюции. 52 (1): 1–7. Дои:10.2307/2410914. JSTOR 2410914. PMID 28568154. Архивировано из оригинал (PDF) на 2019-06-08. Получено 2016-07-08.
- ^ Clutton-Brock TH (май 1989 г.). «Системы спаривания млекопитающих». Труды Лондонского королевского общества. Серия B, Биологические науки. 236 (1285): 339–72. Bibcode:1989RSPSB.236..339C. Дои:10.1098 / rspb.1989.0027. PMID 2567517. S2CID 84780662.
- ^ Boness DJ, Bowen D, Buhleier BM, Marshall GJ (2006). «Тактика спаривания и система спаривания водно-ластоногих: морской тюлень, Phoca vitulina". Поведенческая экология и социобиология. 61: 119–130. Дои:10.1007 / s00265-006-0242-9. S2CID 25266746.
- ^ Клопфер PH (1981). «Истоки родительской заботы». В Губернике DJ (ред.). Родительская забота о млекопитающих. Нью-Йорк: Пленум Пресс. ISBN 978-1-4613-3150-6. OCLC 913709574.
- ^ Мурти Р., Бирман Г., Браун С., Брайант К., Чинн Р., Хьюлетт А. и др. (Май 2015 г.). «Животные в учреждениях здравоохранения: рекомендации по минимизации потенциальных рисков» (PDF). Инфекционный контроль и больничная эпидемиология. 36 (5): 495–516. Дои:10.1017 / ice.2015.15. PMID 25998315.
- ^ Гуманное общество Соединенных Штатов. «Статистика владения домашними животными в США». Получено 27 апреля 2012.
- ^ USDA. "Профиль кролиководства США" (PDF). Архивировано из оригинал (PDF) 7 августа 2019 г.. Получено 10 июля 2013.
- ^ Маккай Р. (26 мая 2013 г.). «Доисторическое наскальное искусство в Дордони». Хранитель. Получено 9 ноября 2016.
- ^ Джонс Дж. (27 июня 2014 г.). «10 лучших портретов животных в искусстве». Хранитель. Получено 24 июн 2016.
- ^ "Охота на оленей в Соединенных Штатах: анализ демографических данных и поведения охотников. Приложение к Национальному обзору рыболовства, охоты и отдыха, связанных с дикой природой 2001-6, 2001 г.". Служба рыболовства и дикой природы (США). Получено 24 июн 2016.
- ^ Шелтон Л. (05.04.2014). "Популярность любительской охоты на свиней растет". Натчез-демократ. Грэмд вид на открытом воздухе. Архивировано из оригинал 12 декабря 2017 г.. Получено 24 июн 2016.
- ^ Нгуен Дж, Уитли Р. (2015). Охота за едой: руководство по сбору урожая, полевой заправке и приготовлению диких животных. F + W Media. С. 6–77. ISBN 978-1-4403-3856-4. Главы об охоте на оленя, кабана, кролика и белки.
- ^ "Скачки". Британская энциклопедия. Архивировано из оригинал 21 декабря 2013 г.. Получено 6 мая 2014.
- ^ Гендеры R (1981). Энциклопедия бега борзых. Книги Пелхэма. ISBN 978-0-7207-1106-6. OCLC 9324926.
- ^ Плюс S (1993). «Роль животных в человеческом обществе». Журнал социальных проблем. 49 (1): 1–9. Дои:10.1111 / j.1540-4560.1993.tb00906.x.
- ^ Фаулер К.Дж. (26 марта 2014 г.). «Топ-10 книг об умных животных». Хранитель. Получено 9 ноября 2016.
- ^ Гэмбл N, Йейтс S (2008). Изучение детской литературы (2-е изд.). Лос-Анджелес: Сейдж. ISBN 978-1-4129-3013-0. OCLC 71285210.
- ^ «Книги для взрослых». Сидящие на тюленях. Получено 9 ноября 2016.
- ^ Патерсон Дж (2013). «Животные в кино и СМИ». Оксфордские библиографии. Дои:10.1093 / obo / 9780199791286-0044.
- ^ Джонс С. (2011). Крупный рогатый скот: история, мифы, искусство. Лондон: Издательство Британского музея. ISBN 978-0-7141-5084-0. OCLC 665137673.
- ^ van Gulik RH. Хаягрива: мантраянский аспект культа лошади в Китае и Японии. Brill Archive. п. 9.
- ^ Грейнджер Р. (24 июня 2012 г.). «Изображение льва в древних и современных религиях». ТРЕВОГА. Архивировано из оригинал 23 сентября 2016 г.. Получено 6 ноября, 2016.
- ^ «Графические детали, карты, инфографика. Подсчет цыплят». Экономист. 27 июля 2011 г.. Получено 6 ноября 2016.
- ^ "Породы крупного рогатого скота СЕГОДНЯ". Крупный рогатый скот сегодня. Cattle-today.com. Получено 6 ноября, 2016.
- ^ Люкефар SD, Cheeke PR. «Стратегии развития кролиководческих проектов в системах натурального хозяйства». Продовольственная и сельскохозяйственная организация. Получено 6 ноября, 2016.
- ^ Пруд РГ (2004). Энциклопедия зоотехники. CRC Press. С. 248–250. ISBN 978-0-8247-5496-9. OCLC 57033325.
- ^ «История кожи». Мур и Джайлз. Получено 10 ноября 2016.
- ^ Браатен А.В. (2005). "Шерсть". В Стиле V (ред.). Энциклопедия одежды и моды. 3. Томсон Гейл. стр.441–443. ISBN 978-0-684-31394-8. OCLC 963977000.
- ^ Quiggle C (осень 2000 г.). «Альпака: древняя роскошь». Трикотажное переплетение: 74–76.
- ^ «Генетические исследования». Фонд здоровья животных. Архивировано из оригинал 12 декабря 2017 г.. Получено 6 ноября, 2016.
- ^ «Разработка лекарств». Animal Research.info. Получено 6 ноября, 2016.
- ^ «Статистика ЕС показывает снижение количества исследований на животных». Кстати об исследованиях. 2013. Получено 6 ноября, 2016.
- ^ Пилчер HR (2003). "Это нокаут". Природа. Дои:10.1038 / news030512-17. Получено 6 ноября, 2016.
- ^ «Поставки и использование приматов в ЕС». Европейская ассоциация биомедицинских исследований. 1996. Архивировано с оригинал на 2012-01-17.
- ^ Карлссон Х.Э., Шапиро С.Дж., Фарах И., Хау Дж. (Август 2004 г.). «Использование приматов в исследованиях: глобальный обзор». Американский журнал приматологии. 63 (4): 225–37. Дои:10.1002 / ajp.20054. PMID 15300710.
- ^ Weatherall D, et al. (2006). Использование нечеловеческих приматов в исследованиях (PDF) (Отчет). Лондон, Великобритания: Академия медицинских наук. Архивировано из оригинал (PDF) 23 марта 2013 г.
- ^ Diamond JM (1997). «Часть 2: Рост и распространение производства продуктов питания». Оружие, микробы и сталь: судьбы человеческих обществ (1-е изд.). Нью-Йорк: W.W. Нортон и компания. ISBN 978-0-393-03891-0. OCLC 35792200.
- ^ Ларсон Дж., Бургер Дж. (Апрель 2013 г.). «Взгляд популяционной генетики на одомашнивание животных» (PDF). Тенденции в генетике. 29 (4): 197–205. Дои:10.1016 / j.tig.2013.01.003. PMID 23415592.
- ^ Зедер М.А. (август 2008 г.). «Одомашнивание и раннее земледелие в Средиземноморском бассейне: истоки, распространение и влияние». Труды Национальной академии наук Соединенных Штатов Америки. 105 (33): 11597–604. Bibcode:2008PNAS..10511597Z. Дои:10.1073 / pnas.0801317105. ЧВК 2575338. PMID 18697943.
- ^ Цена E (2008 г.). Принципы и применения поведения домашних животных: вводный текст. Сакраменто: Издательство Кембриджского университета. ISBN 978-1-84593-398-2. OCLC 226038028.
- ^ Таупиц Дж, Веска М (2009). Химбриды - химеры и гибриды в сравнительных европейских и международных исследованиях. Гейдельберг: Springer. п. 13. ISBN 978-3-540-93869-9. OCLC 495479133.
- ^ Камеры С.М., Файн С.Р., Фацио Б., Амарал М. (2012). «Отчет о систематике североамериканских волков на основе морфологического и генетического анализа». Фауна Северной Америки. 77: 2. Дои:10.3996 / nafa.77.0001.
- ^ ван Вуур Т. (2005). Возвращение зубров - история, морфология и экология вымершего дикого быка. Pensoft Publishers. ISBN 978-954-642-235-4. OCLC 940879282.
- ^ Муни HA, Cleland EE (май 2001 г.). «Эволюционное влияние инвазивных видов». Труды Национальной академии наук Соединенных Штатов Америки. 98 (10): 5446–51. Bibcode:2001ПНАС ... 98.5446М. Дои:10.1073 / pnas.091093398. ЧВК 33232. PMID 11344292.
- ^ Ле Ру JJ, Foxcroft LC, Herbst M, MacFadyen S (январь 2015 г.). «Генетический анализ показывает низкий уровень гибридизации африканских диких кошек (Felis silvestris lybica) и домашних кошек (F. s. Catus) в Южной Африке».. Экология и эволюция. 5 (2): 288–99. Дои:10.1002 / ece3.1275. ЧВК 4314262. PMID 25691958.
- ^ Уилсон А (2003). Отчет о состоянии лесов Австралии. п. 107.
- ^ Раймер Дж. М., Симберлофф Д. (ноябрь 1996 г.). «Вымирание путем гибридизации и интрогрессии». Ежегодный обзор экологии и систематики. 27: 83–109. Дои:10.1146 / annurev.ecolsys.27.1.83.
- ^ Поттс Б.М. (2001). Barbour RC, Hingston AB (ред.). Генетическое загрязнение от сельскохозяйственных лесов с использованием видов и гибридов эвкалиптов: отчет для Совместной программы агролесоводства RIRDC / L & WA / FWPRDC. Австралийская корпорация сельских промышленных исследований и разработок. ISBN 978-0-642-58336-9. OCLC 48794104.
- ^ а б Дирзо Р., Янг Х.С., Галетти М., Себальос Дж., Исаак Нью-Джерси, Коллен Б. (июль 2014 г.). «Дефауна в антропоцене» (PDF). Наука. 345 (6195): 401–6. Bibcode:2014Наука ... 345..401D. Дои:10.1126 / science.1251817. PMID 25061202. S2CID 206555761.
- ^ Primack R (2014). Основы природоохранной биологии (6 изд.). Сандерленд, Массачусетс: Издатели Sinauer Associates, Inc. С. 217–245. ISBN 978-1-60535-289-3. OCLC 876140621.
- ^ Vignieri S (июль 2014 г.). «Исчезающая фауна. Введение». Наука. 345 (6195): 392–5. Bibcode:2014Наука ... 345..392В. Дои:10.1126 / science.345.6195.392. PMID 25061199.
- ^ Берни Д.А., Фланнери Т.Ф. (июль 2005 г.). «Пятьдесят тысячелетий катастрофических исчезновений после контакта с людьми» (PDF). Тенденции в экологии и эволюции. 20 (7): 395–401. Дои:10.1016 / j.tree.2005.04.022. PMID 16701402. Архивировано 10 июня 2010 года.CS1 maint: BOT: статус исходного URL-адреса неизвестен (связь)
- ^ Даймонд Дж (1984). «Исторические исчезновения: Розеттский камень для понимания доисторических исчезновений». В Martin PS, Klein RG (ред.). Четвертичное вымирание: доисторическая революция. Тусон: Университет Аризоны Press. С. 824–862. ISBN 978-0-8165-1100-6. OCLC 10301944.
- ^ Уоттс Дж. (6 мая 2019 г.). «Человеческое общество находится под острой угрозой из-за утраты естественной жизни на Земле». Хранитель. Получено 1 июля, 2019.
- ^ McGrath M (6 мая 2019 г.). «Природный кризис: человечество угрожает исчезновению 1 млн видов.'". BBC. Получено 1 июля, 2019.
- ^ Бар-Он, Йинон М; Филлипс, Роб; Майло, Рон (2018). «Распределение биомассы на Земле». Труды Национальной академии наук. 115 (25): 6506–6511. Дои:10.1073 / pnas.1711842115. ЧВК 6016768. PMID 29784790.
- ^ Main D (22 ноября 2013 г.). «7 знаковых животных, которых люди стремятся к исчезновению». Живая наука.
- ^ Platt JR (25 октября 2011 г.). «Браконьеры доводят яванского носорога до исчезновения во Вьетнаме». Scientific American. Архивировано из оригинал 6 апреля 2015 г.
- ^ Эстрада А., Гарбер П.А., Риландс А.Б., Роос С., Фернандес-Дуке Е., Ди Фьоре А. и др. (Январь 2017 г.). «Надвигающийся кризис вымирания приматов в мире: почему приматы имеют значение». Достижения науки. 3 (1): e1600946. Bibcode:2017SciA .... 3E0946E. Дои:10.1126 / sciadv.1600946. ЧВК 5242557. PMID 28116351.
- ^ Флетчер М (31 января 2015 г.). «Панголины: почему этому милому доисторическому млекопитающему грозит исчезновение». Телеграф.
- ^ Кэррингтон Д. (8 декабря 2016 г.). «Жирафы находятся на грани исчезновения после разрушительного упадка, - предупреждают эксперты». Хранитель.
- ^ Гринфилд, Патрик (9 сентября 2020 г.). «Люди эксплуатируют и разрушают природу в беспрецедентных масштабах - доклад». Хранитель. Получено 13 октября, 2020.
- ^ Маккарти, Доннахад (1 октября 2020 г.). «Ужасающие потери диких животных показывают, что конец вымирания начался - но еще не поздно что-то изменить». Независимый. Получено 13 октября, 2020.
- ^ Pennisi E (18 октября 2016 г.). «Люди до исчезновения охотятся на приматов, летучих мышей и других млекопитающих». Наука. Получено 3 февраля 2017.
- ^ Ripple WJ, Abernethy K, Betts MG, Chapron G, Dirzo R, Galetti M и др. (Октябрь 2016 г.). «Охота на диких животных и угроза исчезновения для млекопитающих в мире». Королевское общество открытой науки. 3 (10): 160498. Bibcode:2016RSOS .... 360498R. Дои:10.1098 / rsos.160498. HDL:1893/24446. ЧВК 5098989. PMID 27853564.
- ^ Уильямс М., Заласевич Дж., Хафф П.К., Швэгерл К., Барноский А.Д., Эллис EC (2015). «Биосфера антропоцена». Обзор антропоцена. 2 (3): 196–219. Дои:10.1177/2053019615591020. S2CID 7771527.
- ^ Морелл V (11 августа 2015 г.). «Мясоеды могут ускорить вымирание видов во всем мире, - предупреждает исследование». Наука. Получено 3 февраля 2017.
- ^ Маховина Б., Фили К.Дж., Ripple WJ (декабрь 2015 г.). «Сохранение биоразнообразия: ключ к сокращению потребления мяса». Наука об окружающей среде в целом. 536: 419–431. Bibcode:2015ScTEn.536..419M. Дои:10.1016 / j.scitotenv.2015.07.022. PMID 26231772.
- ^ Редфорд KH (1992). «Пустой лес» (PDF). Бионаука. 42 (6): 412–422. Дои:10.2307/1311860. JSTOR 1311860.
- ^ Перес CA, Nascimento HS (2006). «Влияние охоты на дичь каяпо юго-восточной Амазонии: последствия для сохранения дикой природы в коренных заповедниках тропических лесов». Эксплуатация человека и сохранение биоразнообразия. Темы биоразнообразия и сохранения. 3. С. 287–313. ISBN 978-1-4020-5283-5. OCLC 207259298.
- ^ Альтрихтер М., Боальо Г. (2004). «Распространение и относительное изобилие пекари в аргентинском Чако: ассоциации с человеческим фактором». Биологическое сохранение. 116 (2): 217–225. Дои:10.1016 / S0006-3207 (03) 00192-7.
- ^ Алверсон Д.Л., Фрибург М.Х., Муравски С.А., Папа Дж. Г. (1996) [1994]. «Прилов морских млекопитающих». Глобальная оценка прилова и выброса рыбы. Рим: Продовольственная и сельскохозяйственная организация Объединенных Наций. ISBN 978-92-5-103555-9. OCLC 31424005.
- ^ Glowka L, Burhenne-Guilmin F, Synge H, McNeely JA, Gündling L (1994). Документ МСОП по экологической политике и законодательству. Путеводитель по Конвенции о биоразнообразии. Международный союз охраны природы. ISBN 978-2-8317-0222-3. OCLC 32201845.
- ^ «О МСОП». Международный союз охраны природы. 2014-12-03. Получено 3 февраля 2017.
- ^ Себальос Г., Эрлих П.Р., Барноски А.Д., Гарсия А., Прингл Р.М., Палмер TM (июнь 2015 г.). «Ускоренная потеря современных видов, вызванная деятельностью человека: вступление в шестое массовое исчезновение». Достижения науки. 1 (5): e1400253. Bibcode:2015SciA .... 1E0253C. Дои:10.1126 / sciadv.1400253. ЧВК 4640606. PMID 26601195.
- ^ Фишер Д.О., Бломберг С.П. (апрель 2011 г.). «Корреляты повторного открытия и обнаружения исчезновения у млекопитающих». Ход работы. Биологические науки. 278 (1708): 1090–7. Дои:10.1098 / rspb.2010.1579. ЧВК 3049027. PMID 20880890.
- ^ Себальос Г., Эрлих А.Х., Эрлих PR (2015). Уничтожение природы: человеческое вымирание птиц и млекопитающих. Балтимор: Издательство Университета Джона Хопкинса. п. 69. ISBN 978-1-4214-1718-9.
- ^ Чжиган Дж, Харрис РБ (2008). "Elaphurus davidianus". Красный список видов, находящихся под угрозой исчезновения МСОП. 2008. Получено 2012-05-20.CS1 maint: ref = harv (связь)
- ^ а б МакКинни ML, Schoch R, Yonavjak L (2013). «Сохранение биологических ресурсов». Экология: системы и решения (5-е изд.). Джонс и Бартлетт Обучение. ISBN 978-1-4496-6139-7. OCLC 777948078.
- ^ Перрин В.Ф., Вюрсиг Б.Г., Thewissen JG (2009). Энциклопедия морских млекопитающих. Академическая пресса. п. 404. ISBN 978-0-12-373553-9. OCLC 455328678.
дальнейшее чтение
- Браун WM (2001). «Естественный отбор компонентов мозга млекопитающих». Тенденции в экологии и эволюции. 16 (9): 471–473. Дои:10.1016 / S0169-5347 (01) 02246-7.
- Яффа К., Тахер Н.А. (2006). "Mammalia Palaestina: Млекопитающие Палестины". Палестинский биологический бюллетень (55): 1–46.
- Маккенна MC, Белл SK (1997). Классификация млекопитающих выше видового уровня. Нью-Йорк: издательство Колумбийского университета. ISBN 978-0-231-11013-6. OCLC 37345734.
- Новак Р.М. (1999). Млекопитающие Уокера в мире (6 изд.). Балтимор: Издательство Университета Джона Хопкинса. ISBN 978-0-8018-5789-8. OCLC 937619124.
- Симпсон Г.Г. (1945). «Принципы классификации и классификации млекопитающих». Бюллетень Американского музея естественной истории. 85: 1–350.
- Мерфи WJ, Eizirik E, O'Brien SJ, Madsen O, Scally M, Douady CJ, et al. (Декабрь 2001 г.). «Разрешение раннего излучения плаценты млекопитающих с использованием байесовской филогенетики». Наука. 294 (5550): 2348–51. Bibcode:2001Научный ... 294.2348M. Дои:10.1126 / science.1067179. PMID 11743200. S2CID 34367609.
- Springer MS, Stanhope MJ, Madsen O, de Jong WW (август 2004 г.). «Молекулы плацентарного дерева консолидируют» (PDF). Тенденции в экологии и эволюции. 19 (8): 430–8. Дои:10.1016 / j.tree.2004.05.006. PMID 16701301.
- Воан Т.А., Райан Дж. М., Капзаплевски Н.Дж. (2000). Маммология (4-е изд.). Форт-Уэрт, Техас: Издательство Сондерс-Колледжа. ISBN 978-0-03-025034-7. OCLC 42285340.
- Кригс Дж.О., Чураков Г., Кифманн М., Джордан У., Брозиус Дж., Шмитц Дж. (Апрель 2006 г.). «Ретропозиционные элементы как архив эволюционной истории плацентарных млекопитающих». PLOS Биология. 4 (4): e91. Дои:10.1371 / journal.pbio.0040091. ЧВК 1395351. PMID 16515367.
- Макдональд Д.В., Норрис С. (2006). Энциклопедия млекопитающих (3-е изд.). Лондон: Brown Reference Group. ISBN 978-0-681-45659-4. OCLC 74900519.
внешняя ссылка
- Biodiversitymapping.org - Все отряды млекопитающих в мире с картами распространения
- Палеоценовые млекопитающие, сайт, посвященный возникновению млекопитающих, paleocene-mammals.de
- Эволюция млекопитающих, краткое введение в древних млекопитающих, enchantedlearning.com
- Атлас млекопитающих Европы EMMA из Societas Europaea Mammalogica, European-mammals.org
- Морские млекопитающие мира - Обзор всех морских млекопитающих, включая описания, как полностью водных, так и полуводных, noaa.gov
- Mammalogy.org Американское общество маммологов было основано в 1919 году с целью содействия изучению млекопитающих, и на этом веб-сайте есть библиотека изображений млекопитающих.
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||







| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||







{kind=link}