Лейшмания - Leishmania
| Лейшмания | |
|---|---|
 | |
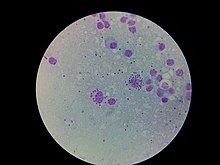
| L. donovani в клетке костного мозга | |
| Научная классификация | |
| Домен: | Эукариоты |
| Тип: | Эвгленозоа |
| Учебный класс: | Кинетопласта |
| Заказ: | Трипаносоматида |
| Род: | Лейшмания Боровский 1898[1] (Росс 1903) |
| Разновидность | |
L. aethiopica | |
Лейшмания /ляʃˈмeɪпяə/ это род из трипаносомы которые несут ответственность за болезнь лейшманиоз.[2][3][4] Они распространяются москиты рода Флеботомус в Старый мир, и рода Луцомия в Новый мир. По крайней мере 93 вида москитов являются подтвержденными или вероятными переносчиками во всем мире.[5] Их основными хозяевами являются позвоночные; Лейшмания обычно заражает даманы, псовые, грызуны, и люди.
История
Первое письменное упоминание о явных симптомах кожный лейшманиоз всплыл в Палеотропы в восточный тексты, относящиеся к 7 веку до н.э. (предположительно переписанные из источников на несколько сотен лет старше, между 1500 и 2000 гг. до н.э.[6]). Из-за того, что лейшманиоз широко и устойчиво распространялся на протяжении всей древности как загадочное заболевание с разнообразными симптомами, лейшманиоз получил различные названия от «белой проказы» до «белой проказы».черная лихорадка ". Некоторые из этих названий предполагают связь с негативными культурными верованиями или мифологией, которые все еще усиливают социальную стигматизацию лейшманиоза сегодня.[7]
Члены древнего род из Лейшмания паразит, Палеолейшмания, были обнаружены в окаменелый мухи начиная с раннего Меловой период,[8] однако возбудитель болезни был обнаружен только в 1901 году в результате одновременного открытия Уильям Буг Лейшман и Чарльз Донован. Они независимо друг от друга визуализировали микроскопических одноклеточных паразитов (позже названных тельцами Лейшмана-Донована), живущих в клетках инфицированных органов человека. Позже паразитический род будет классифицирован как трипаносоматид простейшие под филогенетический обозначение Leishmania donovani. С тех пор несколько видов были классифицированы и сгруппированы в два основных подрода, т.е. Лейшмания Вианния (обычно находится в Неотропы ) или же Лейшмания Лейшмания (обычно находится в Палеотропы, за основным исключением L. mexicana подгруппа).
Эпидемиология
Лейшмания в настоящее время поражает 6 миллионов человек в 98 странах. Ежегодно возникает около 0,9–1,6 миллиона новых случаев, и известно, что 21 вид вызывает заболевания у людей.
Структура
Лейшмания виды одноклеточные эукариоты имея четко определенный ядро и другие клеточные органеллы, включая кинетопласты и жгутики. В зависимости от стадии своего жизненного цикла они существуют в двух структурных вариантах:[9][10]
- В амастигота Форма встречается в мононуклеарных фагоцитах и кровеносных системах человека. Это внутриклеточная неподвижная форма, лишенная внешних жгутиков. Короткий жгутик на переднем конце встроен, но не выступает наружу. Он имеет овальную форму и имеет длину 3–6 мкм и ширину 1–3 мкм. Кинетопласт и базальное тело лежат ближе к переднему концу.
- В промастигота форма находится в пищеварительный тракт москитов. Это внеклеточная и подвижная форма. Он значительно больше и сильно удлинен, его длина составляет 15-30 мкм, а ширина - 5 мкм. Он имеет веретенообразную форму, сужается с обоих концов. Длинный жгутик (примерно равной длине тела) на переднем конце выступает наружу. В центре лежит ядро, а перед ним - кинетопласт и базальное тело.
Эволюция
Детали эволюции этого рода обсуждаются, но Лейшмания очевидно произошли от наследственной линии трипаносом. Самая старая родословная - это Bodonidae, с последующим Trypanosoma brucei, причем последний приурочен к африканскому континенту. Trypanosoma cruzi группы с трипаносомы от летучих мышей, южноамериканских млекопитающих и кенгуру предполагают происхождение в Южном полушарии. Эти клады имеют лишь отдаленное родство.
Остальные клады в этом дереве Бластокритидии, Герпетомонады, и Фитомонады. Четыре рода Лептомонада, Критидия, Лейшмания, и Эндотрипан образуют концевые ветви, предполагая относительно недавнее происхождение. Некоторые из этих родов могут быть полифилетическими и могут нуждаться в дальнейшем делении.[11]
Истоки рода Лейшмания сам непонятен.[12][13] Одна теория предполагает африканское происхождение с миграцией в Америку. Другой предлагает миграцию из Америки в Старый мир через Берингов пролив Сухопутный мост около 15 миллионов лет назад. Третья теория предлагает Палеарктика источник.[14] Такие миграции повлекут за собой последующую миграцию переносчиков и резервуаров или последовательные адаптации на этом пути. Более поздняя миграция - это миграция L. infantum из стран Средиземноморья в Латинская Америка (известный как Л. чагаси), поскольку европейская колонизация Новый мир, где паразиты переняли свой нынешний Новый Свет векторов в их соответствующих экосистемах.[15] Это очевидная причина эпидемий. Одна из недавних эпидемий в Новом Свете коснулась песцов в США.[16]
Хотя предполагалось, что Лейшмания мог развиться в Неотропы.[17] это, вероятно, верно для видов, относящихся к подродам Вианния и Эндотрипан. Однако есть свидетельства того, что первичная эволюция подродов Лейшмания и Сауролейшмания это Старый Свет. В то время как Mundinia виды кажутся более универсальными в своей эволюции. Одна из теорий состоит в том, что разные линии были изолированы географически в разные периоды, и именно это привело к возникновению этого эволюционного мозаицизма. Но несомненно, что Leishmaniinae - монофилетическая группа.
Анализ большого набора данных показывает, что Лейшмания возникла от 90 до 100 миллионов лет назад в Гондвана.[18] Виды, заражающие рептилий, произошли от кладов млекопитающих.
Сауролейшмания виды были первоначально определены на основании того, что они инфицированы рептилии (ящерицы ) скорее, чем млекопитающие. На основании молекулярных данных они были переведены в статус подрода в пределах Лейшмания. Этот подрод, вероятно, произошел от группы, первоначально инфицировавшей млекопитающих.[19]
Таксономия
В этом роде признано 53 вида. Статус некоторых из них оспаривается, поэтому окончательное число может отличаться. По крайней мере 20 видов заражают людей. Чтобы усложнить задачу, гибриды могут быть вовлечены, как сообщалось в Бразилии о гибриде между Leishmania (V.) guyanensis и Лейшмания (В.) шави шави.[20]
В настоящее время род делится на 4 подрода: Лейшмания, Сауролейшмания, Mundinia и Вианния. Разделение на два подрода (Лейшмания и Вианния) был сделан Лейнсоном и Шоу в 1987 году на основании их расположения в кишечнике насекомых. Виды в Вианния подрод развиваются в задней кишке: L. (V.) braziliensis был предложен как типовой вид для этого подрода. Это разделение подтверждено всеми последующими исследованиями. Шоу, Камарго и Тейшейра создали подрод Mundinia при пересмотре Leishmaniinae в 2016 г.[21]
Эндотрипан тесно связан с Лейшмания. Немного Эндотипанум виды уникальны тем, что заражают эритроциты своих хозяев (ленивцев). Все виды обитают в Центральной и Южной Америке.[22] E. colombiensis инфекции были обнаружены у человека.
Сауролейшмания был первоначально описан Ranquein 1973 как отдельный род, но молекулярные исследования показывают, что это фактически подрод, а не отдельный род.
Предлагаемое разделение Лейшмания в Euleishmania и Паралейшмания группы в 2000 году подчеркнули глубокую филогенную дистанцию между паразитами, некоторые из которых были названы Лейшмания разновидность.[23] В Euleishmania включены виды, в настоящее время относящиеся к подродам Лейшмания, Сауролейшмания, Mundinia и Вианния. Предлагаемый Паралейшмания включены виды Эндотипан, Лейшмамния-L. colomubensis, Л. Эррери, L. hertigiи L. deanei и L. equatorensis. В недавнем пересмотре этим видам был присвоен другой родовой статус.
Четыре подрода Лейшмания теперь признаны - Лейшмания, Сауролейшмания, Вианния и Mundinia (в L. enriettii сложный). Род Эндотрипан и Порцизия принадлежат к Паралейшмания.
Есть четыре Mundinia разновидность - L. (Mundinia) enriettii, L. (Mundinia) martiniquensis, L. (Mundinia) макроподум, и L. (Mundinia) orientalis, который находится в Таиланде.[24]
Л. арчибальдиконкретный статус неурегулирован, но он тесно связан с L. donovani.
Л. Эррери принадлежит к роду Эндотипанум а не Лейшмания.
L. donovani и L. infantum тесно связаны.
Примечания
Селенофермент Селтрип кажется уникальным для этого заказа.[25] Он был исключен из подрода Вианния.
L. deanei и L. hertigi, оба из которых заражают дикобразов, были перемещены в род Порцизия.
Классификация
Подрод Лейшмания Росс, 1903 г., сенсю Сафьянова, 1982 г.
- Leishmania aethiopica Брэй, Эшфорд и Брэй, 1973
- Leishmania amazonensis Лейнсон и Шоу, 1972 г. (включает Гарнхами Скорца и др., 1979)
- Leishmania arabica Питерс, Эльбихари и Эванс, 1986
- Leishmania aristidesi Лейнсон и Шоу, 1979
- Leishmania donovani (Лаверан и Менил, 1903 г.)
- Leishmania forattinii Йошида, Куба, Пачеко, Куполильо, Таварес, Мачадо, Хомен и Гримальди, 1993 г.
- Leishmania gerbilli Ван, Цюй и Гуань, 1964 год.
- Leishmania infantum Николь, 1908 год (подвиды чагаси Кунья и Шагас, 1937 год.)
- Leishmania killicki Риу, Ламотт и Пратлонг, 1986 г.
- Leishmania major Якимов и Шохор, 1914 г.
- Leishmania mexicana Бьяджи, 1953 г.
- Leishmania pifanoi Медина и Ромеро, 1959 год.
- Leishmania tropica (Райт, 1903 г.)
- Leishmania turanica Стрелкова, Шурхал, Келлина, Елисеев, Эванс, Питерс, Чепмен, Ле Бланк и Ван Эйс, 1990 г.
- Leishmania venezeulensis Бонфанте-Гарридо, 1980 г.
- Leishmania waltoni Шоу, Пратлонг и Дедет 2015
Подрод Mundinia Шоу, Камарго и Тейшейра 2016
- Leishmania enriettiiМунис и Медина, 1948 г.
- Leishmania macropodum Барратт, Кауфер, Питерс, Крейг, Лоуренс, Робертс, Ли, МакОлифф, Старк, Эллис, 2017
- Leishmania martiniquensis Десбуа, Пратлонг, Квист и Дедет, 2014 г.[26]
- Leishmania orientalis Джарияпан, Дарунтум, Джайвонг, Чанмол, Интахан, Сор-Суван, Сириясатиен, Сомбун, Бейтс, Бейтс, 2018 г.
Подрод Сауролейшмания Ранк, 1973 г., сенсю Сафьянова, 1982 г.
- Leishmania adleri Хайш, 1958 год.
- Leishmania agamae Дэвид, 1929 год.
- Leishmania ceramodactyli Адлер и Теодор, 1929 г.
- Leishmania gulikae Овезмухаммедов, Сафьянова, 1987 г.
- Leishmania gymnodactyli † Ходукин и Софиев, 1940 г.
- Leishmania helioscopi † Чодукин и Софиев, 1940
- Leishmania hemidactyli Маки, Гупта и Сваминатх, 1923 г.
- Leishmania hoogstraali Макмиллан, 1965 год.
- Leishmania nicollei Чодукин и Софифф, 1940
- Leishmania platycephala Телфорд, 2009 г.
- Leishmania phrynocephali Чодукин и Софифф, 1940
- Leishmania senegalensis Ранк, 1973
- Leishmania sofieffi ↑ Марков, Лукина и Маркова, 1964 г.
- Leishmania tarentolae Веньон, 1921 г.
- Leishmania zmeevi ↑ Андручко и Марков 1955
- Leishmania zuckermani Паперна, Булар, Геринг-Хагенбек и Ландау, 2001 г.
↑ Виды, описанные как Сауролейшмания. Их развитие не похоже на развитие других представителей подрода, поэтому их таксономическое положение сомнительно.
Подрод Вианния Лейнсон и Шоу 1987
- Leishmania braziliensis Вианна, 1911 г.
- Leishmania guyanensis Флок, 1954 год.
- Leishmania lainsoni Сильвейра, Шоу, Брага и Исикава, 1987 год.
- Leishmania lindenbergi Сильвейра, Шоу, Брага и Исикава, 1987 год.
- Leishmania naiffi Лейнсон и Шоу, 1989
- Leishmania panamensis Лейнсон и Шоу, 1972 г.
- Leishmania peruviana Велес, 1913 г.
- Лейшмания шави Лейнсон, Брага и де Соуза, 1989
- Leishmania utingensis Брага, Лайнсон, Исикава и Шоу 2003
Родственные роды
Отношения между этими родами и Лейшмания в настоящее время неясно, но они тесно связаны.
Род Эндотрипан
- Endotrypanum colombiensis (Крейцер, Корредор, Гримальди, Грогль, Роутон, Янг, Моралес, МакМахон-Пратт, Гусман и Теш, 1991)
- Endotrypanum equatorensis (Гримальди, Крейцер, Хасигучи, Гомет, Мимори и Теш, 1992)
- Endotrypanum herreri (Зеледон, Понсе и Мурильо, 1979)
- Endotrypanum monterogeii Шоу, 1969 год.
- Endotrypanum schaudinni Мениль и Бримон, 1908 г.
Род Новимонас Костыгов и Юрченко 2016
- Новимонас эсмеральдас Вотыпка, Костыгов, Маслов и Лукеш 2016
Род Порцизия Шоу, Камарго и Тейшейра 2016
- Porcisia deanei (Лейнсон и Шоу 1977)
- Porcisia hertigi (Herrer 1971)
Род Зелония Шоу, Камарго и Тейшейра 2016
- Зелония австралийская Барратт, Кауфер, Питерс, Крейг, Лоуренс, Робертс, Ли, Маколифф, Старк, Эллис 2017
- Zelonia costaricensis (Юрченко, Лукес, Йирку, Зеледон, Маслов, 2006)
Биохимия и клеточная биология
Биохимия и клеточная биология Лейшмания похож на другие кинетопластиды. У них одни и те же основные морфологические признаки, в том числе один жгутик который имеет впячивание, карман жгутика, в его основании, кинетопласт, который находится в сингле митохондрия, и субпеликулярный массив микротрубочек, составляющих основную часть цитоскелет.
Липофосфогликановое покрытие
Лейшмания обладает липофосфогликан нанесите покрытие на внешнюю часть камеры. Липофосфогликан вызывает толл-подобный рецептор 2, сигнальный рецептор, участвующий в запуске врожденный иммунный ответ у млекопитающих.
Точная структура липофосфогликана варьируется в зависимости от вида и жизненный цикл стадия паразита. Гликановый компонент особенно вариабелен, и различные варианты липофосфогликана могут использоваться в качестве молекулярный маркер для разных стадий жизненного цикла. Лектины, группа растение белки которые связывают различные гликаны, часто используются для обнаружения этих вариантов липофосфогликанов. Например, арахисовый агглютинин связывает конкретный липофосфогликан, обнаруженный на поверхности инфекционной формы L. major.
Липофосфогликан используется паразитом для обеспечения своего выживания в организме хозяина, а механизмы, с помощью которых паразит делает это, сосредоточены на модуляции иммунного ответа хозяина. Это жизненно важно, поскольку Лейшмания паразиты живут внутри макрофаги и необходимо предотвратить их убийство макрофагами. Липофосфогликан играет роль в сопротивлении система комплемента, подавляя окислительный взрыв ответ, вызывающий воспаление ответ и предотвращение естественные Т-клетки-киллеры признавая, что макрофаг инфицирован Лейшмания паразит.
| Тип | Возбудитель | Место расположения |
|---|---|---|
| Кожный лейшманиоз (локальные и диффузные) инфекции проявляются в виде явных кожных реакций. | Самым распространенным является Восточная рана (вызвано видами Старого Света L. major, L. tropica, и L. aethiopica ). В Новом Свете наиболее частыми виновниками являются L. mexicana. | Кожные инфекции чаще всего встречаются при Афганистан, Бразилия, Иран, Перу, Саудовская Аравия и Сирия. |
| Кожно-слизистый лейшманиоз инфекции начинаются как реакция на укус и могут пройти метастаз в слизистую оболочку и стать фатальным. | L. braziliensis | Кожно-слизистые инфекции чаще всего встречаются при Боливия, Бразилия и Перу. Кожно-слизистые инфекции также встречаются в Карамай, Китай Синьцзян-Уйгурский автономный район. |
| Висцеральный лейшманиоз инфекции часто распознаются по лихорадке, отеку печени и селезенки и анемия. Они известны под многими местными названиями, наиболее распространенным из которых, вероятно, является кала азар,[27][28] | Вызывается исключительно видами L. donovani сложный (L. donovani, L. infantum син. Л. чагаси).[2] | Встречается в тропических и субтропических районах всех континентов, кроме Австралия, висцеральные инфекции чаще всего встречаются у Бангладеш, Бразилия, Индия, Непал, и Судан.[2] Висцеральный лейшманиоз также встречается в некоторых частях Китая, таких как провинция Сычуань, провинция Ганьсу и Синьцзян-Уйгурский автономный район. |
Внутриклеточный механизм заражения
Во избежание разрушения иммунная система и процветать, Лейшмания «прячется» внутри клеток хозяина. Такое расположение позволяет избежать воздействия гуморальный иммунный ответ (поскольку патоген безопасно находится внутри клетки и вне открытого кровотока), и, кроме того, он может предотвратить уничтожение иммунной системой своего хозяина посредством неопасных поверхностных сигналов, которые препятствуют апоптоз. Основные типы клеток Лейшмания инфильтраты фагоцитозный клетки, такие как нейтрофилы и макрофаги.[29]
Обычно фагоцитозная иммунная клетка, такая как макрофаг, поглощает патоген в закрытом помещении. эндосома а затем заполните эту эндосому ферментами, которые переваривают патоген. Однако в случае Лейшманияэти ферменты не действуют, позволяя паразиту быстро размножаться. Этот беспрепятственный рост паразитов в конечном итоге подавляет макрофаги хозяина или другие иммунные клетки, вызывая их гибель.[30]
Передано москита, то простейшие паразиты из L. major может переключить стратегию первой иммунной защиты с еды / воспаления / убийства на еду / отсутствие воспаления / отсутствие убийства своего хозяина фагоцит и развратили его для своей выгоды.[нужна цитата ] Они неукоснительно используют охотно фагоцитирующие полиморфноядерные нейтрофильные гранулоциты (PMN) как хитрое убежище, где они размножаться не распознаются иммунной системой и входят в долгоживущие макрофаги установить «скрытый» инфекционное заболевание.[нужна цитата ]
Поглощение и выживание

На микробный При инфекции PMN перемещаются из кровотока через эндотелиальный слой сосудов к участку инфицированной ткани (кожная ткань после укуса мухи). Они немедленно инициируют первый иммунный ответ и фагоцитируют захватчика, распознавая чужеродные и активируя поверхности на паразите. Активированный секрет PMN хемокины, Ил-8 в частности, чтобы привлечь больше гранулоциты и стимулируют фагоцитоз. Дальше, L. major увеличивает секрецию IL-8 PMN. Этот механизм наблюдается при заражении другими облигатные внутриклеточные паразиты, также. Для подобных микробов существует множество механизмов внутриклеточного выживания. Удивительно, но совместная инъекция апоптотических и жизнеспособных патогенов вызывает гораздо более скоротечное течение болезни, чем инъекции только жизнеспособных паразитов. Когда противовоспалительный сигнал фосфатидилсерин обычно обнаруживается на апоптотических клетках, обнажается на поверхности мертвых паразитов, L. major выключает окислительный взрыв, тем самым предотвращая гибель и разложение жизнеспособного патогена.
В случае Лейшмания, потомство не генерируется в PMNs, но таким образом они могут выжить и сохраняться не запутанными в первичном очаге инфекции. Формы промастигот также выпускают Лейшмания хемотаксический фактор (LCF) для активного рекрутирования нейтрофилов, но не других лейкоциты, например моноциты или же NK-клетки. Кроме того, производство интерферон гамма (IFNγ) -индуцируемый белок 10 (IP10) PMN блокируется при участии Лейшмания, что включает в себя отключение воспалительного и защитного иммунного ответа NK и Клетка Th1 набор персонала. Патогены остаются жизнеспособными во время фагоцитоза, так как их первичные хозяева, PMN, раскрывают молекулярный паттерн, связанный с апоптотическими клетками (ACAMP), сигнализируя «нет патогена».
Настойчивость и влечение
Продолжительность жизни нейтрофильные гранулоциты довольно коротко. Они циркулируют в кровоток примерно от 6 до 10 часов после отъезда Костный мозг, после чего они подвергаются спонтанному апоптоз. Сообщалось, что микробные патогены влияют на клеточный апоптоз разными способами. Очевидно из-за торможения каспаза 3-активация, L. major может вызвать задержку апоптоза нейтрофилов и продлить их жизнь как минимум на 2–3 дня. Факт увеличения продолжительности жизни очень благоприятен для развития инфекции, потому что конечными клетками-хозяевами для этих паразитов являются макрофаги, которые обычно мигрируют в места заражения в течение двух или трех дней. Возбудители не дрянные; вместо этого они берут на себя управление первичным очагом заражения. Они индуцируют продукцию PMN хемокинов MIP-1α и MIP-1β (воспалительный белок макрофагов ) для набора макрофагов.[31]
Теория тихого фагоцитоза
Чтобы сохранить целостность окружающей ткани от токсичный компоненты клетки и протеолитический ферменты содержащиеся в нейтрофилах, апоптотические PMN молча выводятся макрофагами. Умирающие PMN выставляют сигнал "съешь меня" фосфатидилсерин который переносится на внешний листок плазматическая мембрана во время апоптоза. Из-за отсроченного апоптоза паразиты, сохраняющиеся в PMN, попадают в макрофаги, используя абсолютно физиологический и нефлогистический процесс. Стратегия этого «тихого фагоцитоза» дает паразиту следующие преимущества:
- Поглощение апоптотических клеток подавляет активность макрофагов, убивающую их, что приводит к выживанию патогенов.
- Патогены внутри PMN не имеют прямого контакта с поверхностью макрофагов рецепторы, потому что они не могут видеть паразита внутри апоптотической клетки. Итак, активация фагоцита для иммунной активации не происходит.
Однако исследования показали, что это маловероятно, поскольку видно, что патогены покидают апоптопные клетки, и нет никаких доказательств поглощения макрофагами этим методом.
Молекулярная биология
Важный аспект Лейшмания простейшие это его гликоконъюгат слой липофосфогликан (СНГ). Он удерживается вместе с якорем фосфоинозитидной мембраны и имеет трехчастную структуру, состоящую из липидного домена, нейтрального гексасахарида и фосфорилированной галактозо-маннозы с концевой нейтралью. У этих паразитов не только развивается постфлеботомусное пищеварение, но и считается, что они необходимы для окислительных всплесков, что позволяет проникать инфекции. Характеристики внутриклеточного пищеварения включают: эндосома слияние с лизосома, высвобождая кислоту гидролазы которые ухудшают ДНК, РНК, белки и углеводы.
Геномика
Геномы четырех Лейшмания разновидность (L. major, L. infantum, L. donovani и L. braziliensis) были секвенированы, выявив более 8300 кодирующих белков и 900 РНК гены. Почти 40% генов, кодирующих белок, относятся к 662 семействам, содержащим от двух до 500 членов. Большинство меньших семейств генов представляют собой тандемные массивы от одного до трех генов, в то время как большие семейства генов часто рассредоточены в тандемных массивах в разных местах. места на протяжении геном. Каждый из 35 или 36 хромосомы организован в небольшое количество кластеров генов от десятков до сотен генов на одной и той же цепи ДНК. Эти кластеры могут быть организованы по типу «голова к голове» (расходящиеся) или «хвост к хвосту» (конвергентные), причем последние часто разделяются тРНК, рРНК и / или мяРНК гены. Транскрипция генов, кодирующих белок, инициируется двунаправленно в дивергентных областях переключения цепей между кластерами генов и расширяет полицистронически через каждый кластер генов перед завершением в области переключения цепи, разделяющей конвергентные кластеры. Лейшмания теломеры обычно являются относительно небольшими и состоят из нескольких повторяющихся последовательностей различных типов. Можно найти доказательства рекомбинации между несколькими различными группами теломер. В L. major и L. infantum геномы содержат всего около 50 копий неактивных вырожденных Инги/ L1Tc-связанные элементы (DIREs), а L. braziliensis также содержит несколько связанных с теломерами подвижных элементов и связанных со сплайсингом ретроэлементов, связанных с лидером. В Лейшмания геномы разделяют консервативный коровой протеом из примерно 6200 генов с родственными трипаносоматидами Trypanosoma brucei и Trypanosoma cruzi, но около 1000 Лейшманияизвестны -специфические гены, которые в основном случайным образом распределены по геному. Относительно немного (около 200) видоспецифичных различий в содержании генов существует между тремя секвенированными Лейшмания геномов, но около 8% генов, по-видимому, эволюционируют с разной скоростью между тремя видами, что указывает на различное давление отбора, которое может быть связано с патологией заболевания. Около 65% генов, кодирующих белок, в настоящее время не имеют функционального назначения.[3]
Лейшмания виды производят несколько разных белки теплового шока. К ним относятся Hsp83, гомолог Hsp90. Регулирующий элемент в 3 'UTR элементов управления Hsp83 перевод Hsp83 чувствительным к температуре способом. Этот регион образует стабильную Структура РНК который плавится при более высоких температурах.[32]
Половое размножение
Репродуктивная система микробного патогена является одним из основных биологических процессов, определяющих экологию микроорганизма и распространение болезни.[33] В 2009 г. Акопянц и др.[34] продемонстрировал, что L. major имеет половой цикл, в том числе мейотический процесс. Формируются гибридные потомки, которые имеют полный геномный набор от обоих родителей. Спаривание происходит только в песчаная муха вектор, а гибриды могут передаваться млекопитающему-хозяину через укус песчаной мухи. В L. braziliensis спаривания в природе преимущественно происходят между родственными особями, что приводит к крайнему инбридингу.[35] Скорость ауткроссинга между разными штаммами Лейшмания в переносчике песчаных мух зависит от частоты сочетанного заражения. Такие случаи ауткроссинга в L. major [34] и L. donovani.[36]
L. infantum продуцирует белки BRCA1 и RAD51, которые взаимодействуют друг с другом, способствуя гомологичной рекомбинационной репарации.[37] Эти белки играют ключевую роль в мейозе. Таким образом, мейотические события обеспечивают адаптивное преимущество эффективной рекомбинационной репарации повреждений ДНК, даже если они не приводят к ауткроссингу.[нужна цитата ]
Смотрите также
Рекомендации
- ^ Шнур, Л. Ф (1989). «О клинических проявлениях и паразитах лейшманиозов Старого Света и Leishmania Tropica, вызывающих висцеральный лейшманиоз». В Харте, Д. Т. (ред.). Лейшманиоз. С. 939–43. Дои:10.1007/978-1-4613-1575-9_119. ISBN 978-1-4612-8862-6.
- ^ а б c Райан К.Дж.; Рэй CG (редакторы) (2004). Шеррис Медицинская микробиология (4-е изд.). Макгроу Хилл. С. 749–54. ISBN 0-8385-8529-9.CS1 maint: несколько имен: список авторов (связь) CS1 maint: дополнительный текст: список авторов (связь)
- ^ а б Myler P; Фазель Н (редакторы) (2008). Лейшмания: после генома. Caister Academic Press. ISBN 978-1-904455-28-8.CS1 maint: несколько имен: список авторов (связь) CS1 maint: дополнительный текст: список авторов (связь)
- ^ Ansari MY, Equbal A, Dikhit MR, Mansuri R, Rana S, Ali V, Sahoo GC, Das P (ноябрь 2015 г.). «Установление корреляции между тестовым анализом In-Silico и In-vitro против Leishmania HGPRT с ингибиторами». Международный журнал биологических макромолекул. 83: 78–96. Дои:10.1016 / j.ijbiomac.2015.11.051. PMID 26616453.
- ^ ВОЗ (2010) Годовой отчет. Женева
- ^ Кокс, Ф. Э. Г (2002). «История паразитологии человека». Обзоры клинической микробиологии. 15 (4): 595–612. Дои:10.1128 / CMR.15.4.595-612.2002. ЧВК 126866. PMID 12364371.
- ^ Яник, М .; и другие. (2004). «Психологические последствия кожного лейшманиоза». Clin Exp Дерматол. 29 (5): 464–467. Дои:10.1111 / j.1365-2230.2004.01605.x. PMID 15347324.
- ^ Пойнар, G (2008). «Lutzomyia adiketis sp. N. (Diptera: Phlebotomidae), переносчик Paleoleishmania neotropicum sp. N. (Kinetoplastida: Trypanosomatidae) в доминиканском янтаре». Векторы паразитов. 1 (1): 2. Дои:10.1186/1756-3305-1-22. ЧВК 2491605. PMID 18627624.
- ^ «Морфология и жизненный цикл». UCLA. Получено 24 января 2014.
- ^ Pulvertaft, RJ; Хойл, GF (1960). «Этапы жизненного цикла Leishmania donovani». Труды Королевского общества тропической медицины и гигиены. 54 (2): 191–6. Дои:10.1016/0035-9203(60)90057-2. PMID 14435316.
- ^ Хьюз, Алабама; Пионтковская, H (2003). «Филогения Trypanosomatidae и Bodonidae (Kinetoplastida) на основе 18S рРНК: доказательства парафилии Трипаносома и шесть других родов ". Мол Биол Эвол. 20 (4): 644–652. Дои:10.1093 / molbev / msg062. PMID 12679543.
- ^ Момен Х, Куполильо Э (2000). «Размышления о происхождении и эволюции рода Leishmania». Mem. Inst. Освальдо Крус. 95 (4): 583–8. Дои:10.1590 / S0074-02762000000400023. PMID 10904419.
- ^ Нойес Х.А., Моррисон Д.А., Шанс М.Л., Эллис Дж.Т. (2000). «Доказательства неотропического происхождения лейшмании». Mem. Inst. Освальдо Крус. 95 (4): 575–8. Дои:10.1590 / S0074-02762000000400021. PMID 10904417.
- ^ Керр С.Ф. (2000). «Палеарктическое происхождение Leishmania». Mem. Inst. Освальдо Крус. 95 (1): 75–80. Дои:10.1590 / S0074-02762000000100011. PMID 10656708.
- ^ Кулс, Катрин; Алам, Мохаммад Захангир; Куполильо, Элиза; Феррейра, Габриэль Эдуардо М .; Mauricio, Isabel L .; Оддоне, Роландо; Фелициангели, М. Дора; Вирт, Тьерри; Майлз, Майкл А .; Шениан, Габриэле; Камхави, Шаден (7 июня 2011 г.). «Сравнительное микросателлитное типирование Leishmania infantum в Новом Свете выявляет низкую гетерогенность среди популяций и ее недавнее происхождение из Старого Света». PLoS забытые тропические болезни. 5 (6): e1155. Дои:10.1371 / journal.pntd.0001155. ЧВК 3110170. PMID 21666787.
- ^ Duprey, Z. H .; Steurer, F.J .; Руни, Дж. А .; Кирхгоф, Л. В .; Джексон, Дж. Э .; Rowton, E.D .; Шанц, П. М. (2006). "Висцеральный лейшманиоз собак, США и Канада, 2000–2003 гг.". Возникающие инфекционные заболевания. 12 (3): 440–446. Дои:10.3201 / eid1203.050811. ЧВК 3291440. PMID 16704782.
- ^ Нойес, штат Гавайи; Arana, BA; Шанс, мл; Maingon, R (1997). "The Leishmania hertigi (Kinetoplastida; Trypanosomatidae) комплекс и ящерица Лейшмания: их классификация и доказательства неотропического происхождения Лейшмания-Эндотрипан клады ". J Eukaryot Microbiol. 44 (5): 511–557. Дои:10.1111 / j.1550-7408.1997.tb05732.x.
- ^ Харкинс, Келли М; Шварц, Рэйчел С; Картрайт, Рид А; Стоун, Энн С. (2016). «Филогеномная реконструкция подтверждает происхождение Leishmania на суперконтиненте». Инфекция, генетика и эволюция. 38: 101–9. Дои:10.1016 / j.meegid.2015.11.030. PMID 26708057.
- ^ Кроан, Дэвид Джи; Моррисон, Дэвид А; Эллис, Джон Т (1997). «Эволюция рода Leishmania выявлена путем сравнения последовательностей генов ДНК и РНК-полимеразы». Молекулярная и биохимическая паразитология. 89 (2): 149–59. Дои:10.1016 / S0166-6851 (97) 00111-4. PMID 9364962.
- ^ Jennings, Y.L .; де Соуза, А. А. А .; Ishikawa, E.A .; Shaw, J .; Lainson, R .; Сильвейра, Ф. (2014). «Фенотипическая характеристика Лейшмания виды вызывая кожный лейшманиоз в нижнем течении Амазонки, западный штат Пара, Бразилия, выявляет предполагаемого гибридного паразита, Лейшмания (Вианния) гайаненсис × Лейшмания (Вианния) шави шави". Паразит. 21: 39. Дои:10.1051 / паразит / 2014039. ЧВК 4118625. PMID 25083790.
- ^ Espinosa, O.A .; Серрано, М.Г .; Camargo, E.P .; M.M.G., Тейшейра; Джей-Джей, Шоу (2016). "Оценка таксономии и номенклатуры трипаносоматид, которые в настоящее время классифицируются как Leishmania '' и '' Endotrypanum". Паразитология. 145: 430–442. Дои:10.1017 / S0031182016002092. PMID 27976601.
- ^ Франко, AM; Гримальди, Дж. Младший (1999). "Характеристика Эндотрипан (Kinetoplastida: Trypanosomatidae), уникальный паразит, поражающий неотропических древесных ленивцев (Edentata) ». Mem Inst Oswaldo Cruz. 94 (2): 261–268. Дои:10.1590 / s0074-02761999000200026. PMID 10224540.
- ^ Момен, H; Куполильо, Э (2000). "Размышления о происхождении и эволюции рода Лейшмания". Mem Inst Oswaldo Cruz. 95 (4): 583–588. Дои:10.1590 / s0074-02762000000400023. PMID 10904419.
- ^ Джарийапан, N; Daroontum, T; Jaiwong, K; Чанмол, Вт; Интахан, Н; Сор-Суван, S; Siriyasatien, P; Somboon, P; Бейтс, доктор медицины; Бейтс, Пенсильвания (2018). "Leishmania (Mundinia) orientalis (Лейшмания восточная) п. sp. (Trypanosomatidae), паразит из Таиланда, ответственный за локализованный кожный лейшманиоз ». Векторы паразитов. 11 (1): 351. Дои:10.1186 / s13071-018-2908-3.
- ^ Бонилла, Мариана; Крулл, Эрика; Иригоин, Флоренсия; Салинас, Густаво; Комини, Марсело А. (2016). «Селенопротеины африканских трипаносом незаменимы для выживания паразитов в организме млекопитающего-хозяина». Молекулярная и биохимическая паразитология. 206 (1–2): 13–19. Дои:10.1016 / j.molbiopara.2016.03.002. PMID 26975431.
- ^ Десбуа, Николь; Пратлонг, Франсин; Квист, Даниэль; Деде, Жан-Пьер (2014). "Лейшмания (Leishmania) martiniquensis п. sp. (Kinetoplastida: Trypanosomatidae), описание паразита, ответственного за кожный лейшманиоз на острове Мартиника (Французская Вест-Индия) ". Паразит. 21: 12. Дои:10.1051 / паразит / 2014011. ISSN 1776-1042. ЧВК 3952653. PMID 24626346.

- ^ Висцеральный лейшмниоз: заболевание В архиве 2005-04-28 на Wayback Machine
- ^ кала-азар В архиве 2009-02-10 на Wayback Machine. Словарь английского языка American Heritage Dictionary
- ^ Ванье-Сантос, Массачусетс; Мартини А; де Соуза В. (август 2002 г.). «Клеточная биология видов Leishmania: вторжение и уклонение». Текущий фармацевтический дизайн. 8 (4): 297–318. Дои:10.2174/1381612023396230. PMID 11860368.
- ^ Пол, Уильям Э. (сентябрь 1993 г.). «Инфекционные болезни и иммунная система». Scientific American. 269 (3): 94–95. Bibcode:1993SciAm.269c..90P. Дои:10.1038 / scientificamerican0993-90. PMID 8211095.
- ^ Laskay T, et al. (2003). «Нейтрофильные гранулоциты - троянские кони для Leishmania major и прочие внутриклеточные микробы? ». Тенденции в микробиологии. 11 (5): 210–4. Дои:10.1016 / S0966-842X (03) 00075-1. PMID 12781523.
- ^ Дэвид, М; Габданк, я; Бен-Давид, М; Зилка, А; Орр, я; Бараш, Д; Шапира, М. (февраль 2010 г.). «Предпочтительная трансляция Hsp83 в Leishmania требует наличия термочувствительного элемента, богатого полипиримидином, в 3 'UTR и включает сканирование 5' UTR». РНК. 16 (2): 364–74. Дои:10.1261 / rna.1874710. ЧВК 2811665. PMID 20040590.
- ^ Rougeron V, De Mees T, Kako Ouraga S, Hide M, Bañuls AL (2010). ""Все, что вы всегда хотели знать о сексе (но боялись спросить) «в Leishmania после двух десятилетий лабораторных и полевых анализов». PLoS Pathog. 6 (8): e1001004. Дои:10.1371 / journal.ppat.1001004. ЧВК 2924324. PMID 20808896.
- ^ а б Акопянц Н.С., Кимблин Н., Секундино Н., Патрик Р., Петерс Н., юрист П., Добсон Д.Е., Беверли С.М., Сакс Д.Л. (апрель 2009 г.). «Демонстрация генетического обмена во время циклического развития Leishmania в векторе песчаных мух». Наука. 324 (5924): 265–8. Bibcode:2009Sci ... 324..265A. Дои:10.1126 / science.1169464. ЧВК 2729066. PMID 19359589.
- ^ Rougeron V, De Mees T, Hide M, Waleckx E, Bermudez H, Arevalo J, Llanos-Cuentas A, Dujardin JC, De Doncker S, Le Ray D, Ayala FJ, Bañuls AL (июнь 2009 г.). «Экстремальный инбридинг у Leishmania braziliensis». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 106 (25): 10224–9. Bibcode:2009PNAS..10610224R. Дои:10.1073 / pnas.0904420106. ЧВК 2700931. PMID 19497885.
- ^ Роджерс МБ, Даунинг Т., Смит Б.А., Имамура Х., Сандерс М., Свободова М., Вольф П., Берриман М., Коттон Дж. А., Смит Д.Ф. (январь 2014 г.). «Геномное подтверждение гибридизации и недавнего инбридинга в изолированной популяции Leishmania». PLoS Genet. 10 (1): e1004092. Дои:10.1371 / journal.pgen.1004092. ЧВК 3894156. PMID 24453988.
- ^ Дженуа М.М., Мукерджи А., Убеда Дж. М., Бюиссон Р., Паке Е., Рой Дж., Плурд М., Куломб И., Уэллетт М., Массон Дж. Ю. (август 2012 г.). «Взаимодействие между BRCA2 и RAD51 для содействия гомологичной рекомбинации у Leishmania infantum». Нуклеиновые кислоты Res. 40 (14): 6570–84. Дои:10.1093 / нар / гкс306. ЧВК 3413117. PMID 22505581.
дальнейшее чтение
- Ван Зандберген, G; Боллинджер, А; Венцель, А; Камхави, S; Voll, R; Клингер, М; Мюллер, А; Holscher, C; Herrmann, M; Мешки, D; Сольбах, Вт; Ласкей, Т. (2006). «Развитие болезни лейшмании зависит от наличия апоптотических промастигот в вирулентном инокуляте». Труды Национальной академии наук. 103 (37): 13837–42. Bibcode:2006ПНАС..10313837В. Дои:10.1073 / pnas.0600843103. ЧВК 1564231. PMID 16945916.
- Шоу, Джеффри Джон (1969). Гемофлагелляты ленивцев . Х. К. Льюис: Лондон.
- Ansari MY, Dikhit MR, Sahoo GC, Das P (2012). «Сравнительное моделирование фермента HGPRT L. donovani и аффинностей связывания различных аналогов GMP». Int J Biol Macromol. 50 (3): 637–49. Дои:10.1016 / j.ijbiomac.2012.01.010. PMID 22327112.
внешняя ссылка
- Международная сеть лейшмании (ILN) содержит основную информацию о болезни и ссылки на многие аспекты болезни и ее переносчиков.
- Список обсуждения (Лейш-Л) также доступен с более чем 600 подписчиками к списку, от молекулярных биологов до работников общественного здравоохранения из многих стран как внутри, так и за пределами эндемичных регионов. Комментарии и вопросы приветствуются.
- KBD: Кинетопластическая биология и болезнь, это веб-сайт, посвященный лейшманиозу, сонной болезни и Болезнь Шагаса (Американский трипаносомоз). Он содержит бесплатный доступ к полному тексту рецензируемых статей по этой тематике. На сайте размещено множество статей, касающихся уникальных кинетопластидных органелл и генетического материала в них.
- Половое размножение у паразитов лейшмании, краткий обзор «научной» статьи
- Сетка мирового сообщества: поиск лекарств от лейшманиоза