Геном - Genome
| Часть серия на |
| Генетика |
|---|
 |
| Ключевые компоненты |
| История и темы |
| Исследование |
| Персонализированная медицина |
| Персонализированная медицина |

В полях молекулярная биология и генетика, а геном это весь генетический материал организма. Это состоит из ДНК (или же РНК в РНК-вирусы ). В геном входят как гены (в кодирующие области ) и некодирующая ДНК,[1] а также митохондриальная ДНК[2] и хлоропластная ДНК. Исследование генома называется геномика.
Происхождение термина
Период, термин геном была создана в 1920 году Ханс Винклер,[3] профессор ботаника на Гамбургский университет, Германия. Оксфордский словарь предполагает, что название представляет собой смесь слов ген и хромосома.[4] Однако см. омики для более подробного обсуждения. Несколько связанных -ome слова уже существовали, например биом и корневище, формируя словарь, в который геном подходит систематически.[5]
Секвенирование и отображение
Последовательность генома - это полный список нуклеотиды (A, C, G и T для геномов ДНК), которые составляют все хромосомы человека или вида. Внутри одного вида у разных особей подавляющее большинство нуклеотидов идентичны, но для понимания генетического разнообразия необходимо секвенирование нескольких особей.

В 1976 г. Уолтер Фирс на Гентский университет (Бельгия) первым установил полную нуклеотидную последовательность вирусного РНК-генома (Бактериофаг MS2 ). В следующем году, Фред Сэнгер завершили первую последовательность ДНК-генома: Фаг Φ-X174, из 5386 пар оснований.[6] Первые полные последовательности генома среди всех трех доменов жизни были выпущены в течение короткого периода в середине 1990-х годов: первым секвенированным бактериальным геномом был геном Haemophilus influenzae, выполненный командой в Институт геномных исследований в 1995 году. Несколько месяцев спустя был завершен первый эукариотический геном с последовательностями 16 хромосом почкующихся дрожжей. Saccharomyces cerevisiae опубликовано в результате усилий европейцев, начатых в середине 1980-х годов. Первая последовательность генома для Археон, Methanococcus jannaschii, был завершен в 1996 году снова Институтом геномных исследований.
Развитие новых технологий сделало секвенирование генома значительно дешевле и проще, а количество полных последовательностей генома быстро растет. В Национальные институты здравоохранения США поддерживает одну из нескольких всеобъемлющих баз данных геномной информации.[7] Среди тысяч завершенных проектов секвенирования генома есть проекты для рис, а мышь, завод Arabidopsis thaliana, то рыба фугу, и бактерии Кишечная палочка. В декабре 2013 года ученые впервые секвенировали всю геном из Неандерталец, вымерший вид люди. Геном был извлечен из кость пальца 130000-летнего неандертальца, найденного в Сибирская пещера.[8][9]
Новые технологии секвенирования, такие как массивное параллельное секвенирование также открыли перспективу персонального секвенирования генома в качестве диагностического инструмента, впервые предложенного Прогнозирующая медицина Manteia. Важным шагом к этой цели стало завершение в 2007 г. полный геном из Джеймс Д. Уотсон, один из соавторов структуры ДНК.[10]
В то время как последовательность генома перечисляет порядок каждой основы ДНК в геноме, карта генома определяет ориентиры. Карта генома менее подробна, чем последовательность генома, и помогает ориентироваться в геноме. В Проект "Геном человека" был организован карта и чтобы последовательность то человеческий геном. Основным шагом в реализации проекта стал выпуск подробной геномной карты Жан Вайссенбах и его команда в Геноскоп в Париже.[11][12]
Контрольный геном последовательности и карты продолжают обновляться, удаляя ошибки и уточняя области высокой аллельной сложности.[13] Снижение стоимости геномного картирования позволило генеалогический сайты, предлагающие это как услугу,[14] в той мере, в какой можно подчинить свой геном краудсорсинг научные усилия, такие как ДНК.ЗЕМЛЯ на Нью-Йоркский центр генома,[15] пример обоих эффект масштаба и из гражданская наука.[16]
Вирусные геномы
Вирусные геномы может состоять из РНК или ДНК. Геномы РНК-вирусы может быть одноцепочечная РНК или же двухцепочечная РНК, и может содержать одну или несколько отдельных молекул РНК (сегменты: одно- или многочастичный геном). ДНК-вирусы могут иметь одноцепочечный или двухцепочечный геном. Большинство геномов ДНК-вирусов состоят из одной линейной молекулы ДНК, но некоторые из них состоят из кольцевой молекулы ДНК.[17]Существуют также вирусные РНК, называемые одноцепочечными РНК: они служат в качестве матрицы для синтеза мРНК.[18] и одноцепочечная РНК: служит матрицей для синтеза ДНК.
Вирусный конверт[19] представляет собой внешний слой мембраны, который вирусные геномы используют для проникновения в клетку-хозяина. Некоторые классы вирусной ДНК и РНК состоят из вирусной оболочки, а некоторые нет.
| Класс / Семья | Конверт? |
|---|---|
| Двухцепочечная ДНК | |
| Аденовирус | Нет |
| Папилломавирус | Нет |
| Полиомавирус | Нет |
| Герпесвирус | да |
| Поксвирус | да |
| Одноцепочечная ДНК | |
| Парвовирус | Нет |
| Двухцепочечная РНК | |
| Реовирус | Нет |
| Одноцепочечная РНК | |
| Пикорнавирус | Нет |
| Коронавирус | да |
| Флавивирус | да |
| Тогавирус | да |
| Одноцепочечная РНК: служит матрицей для синтеза мРНК | |
| Филовирус | да |
| Ортомиксовирус | да |
| Парамиксовирус | да |
| Рабдовирус | да |
| Одноцепочечная РНК: служит матрицей для синтеза ДНК | |
| Ретровирус | да |
Прокариотические геномы
Прокариоты и эукариоты имеют геномы ДНК. Археи и большинство бактерий имеют один круговая хромосома,[20] однако некоторые виды бактерий имеют линейные или множественные хромосомы.[21][22] Если ДНК реплицируется быстрее, чем делятся бактериальные клетки, несколько копий хромосомы могут присутствовать в одной клетке, а если клетки делятся быстрее, чем ДНК может быть реплицирована, множественная репликация хромосомы инициируется до того, как произойдет деление, позволяя дочерним клеткам наследовать полные геномы и уже частично реплицированные хромосомы. У большинства прокариот в геномах очень мало повторяющейся ДНК.[23] Однако некоторые симбиотические бактерии (например. Serratia symbiotica ) имеют уменьшенные геномы и высокую долю псевдогенов: только ~ 40% их ДНК кодирует белки.[24][25]
У некоторых бактерий есть вспомогательный генетический материал, также являющийся частью их генома, который передается в плазмиды. Для этого слово геном не следует использовать как синоним хромосома.
Эукариотические геномы
Геномы эукариот состоят из одной или нескольких линейных хромосом ДНК. Количество хромосом широко варьируется от Джек-джемпер муравьи и бесполый немотод,[26] у каждого есть только одна пара, чтобы виды папоротников что имеет 720 пар.[27] Типичная человеческая клетка имеет две копии каждой из 22 аутосомы, по одному унаследованному от каждого родителя, плюс два половые хромосомы, делая его диплоидным. Гаметы, такие как яйцеклетки, сперма, споры и пыльца, являются гаплоидными, что означает, что они несут только одну копию каждой хромосомы.
Помимо хромосом в ядре, органеллы, такие как хлоропласты и митохондрии имеют собственную ДНК. Иногда говорят, что митохондрии имеют собственный геном, который часто называют "митохондриальный геном ". ДНК, обнаруженная в хлоропласте, может быть обозначена как"пластом ". Подобно бактериям, из которых они произошли, митохондрии и хлоропласты имеют кольцевую хромосому.
В отличие от прокариот, эукариоты имеют экзон-интронную организацию генов, кодирующих белок, и различное количество повторяющейся ДНК. У млекопитающих и растений большая часть генома состоит из повторяющейся ДНК.[28]
Кодирующие последовательности
Последовательности ДНК, которые несут инструкции по созданию белков, называются кодирующими последовательностями. Доля генома, занятая кодирующими последовательностями, широко варьируется. Геном большего размера не обязательно содержит больше генов, и доля неповторяющейся ДНК уменьшается вместе с увеличением размера генома у сложных эукариот.[28]

Некодирующие последовательности
Некодирующие последовательности включают интроны, последовательности для некодирующих РНК, регуляторных областей и повторяющейся ДНК. Некодирующие последовательности составляют 98% генома человека. В геноме есть две категории повторяющейся ДНК: тандем повторяет и вкрапленные повторы.[29]
Тандем повторяет
Короткие некодирующие последовательности, повторяющиеся от головы к хвосту, называются тандем повторяет. Микросателлитные повторы состоят из 2-5 пар оснований, а минисателлитные повторы составляют 30-35 п.н. Тандемные повторы составляют около 4% генома человека и 9% генома плодовой мушки.[30] Тандемные повторы могут быть функциональными. Например, теломеры состоят из тандемного повтора TTAGGG у млекопитающих и играют важную роль в защите концов хромосомы.
В других случаях увеличение количества тандемных повторов в экзонах или интронах может вызвать болезнь.[31] Например, ген хантингтина человека обычно содержит 6–29 тандемных повторов нуклеотидов CAG (кодирующих полиглутаминовый тракт). Расширение до более чем 36 повторов приводит к болезнь Хантингтона, нейродегенеративное заболевание. Известно, что двадцать заболеваний человека возникают в результате аналогичных тандемных повторений в различных генах. Механизм, с помощью которого белки с расширенными полигулатаминовыми путями вызывают гибель нейронов, полностью не изучен. Одна из возможностей заключается в том, что белки не могут правильно складываться и избегать деградации, вместо этого они накапливаются в агрегатах, которые также секвестрируют важные факторы транскрипции, тем самым изменяя экспрессию генов.[31]
Тандемные повторы обычно вызываются проскальзыванием во время репликации, неравным кроссинговером и преобразованием генов.[32]
Переносные элементы
Мобильные элементы (TE) - это последовательности ДНК с определенной структурой, которые способны изменять свое положение в геноме.[30][23][33] ТЕ классифицируются как ТЕ класса I, которые реплицируются с помощью механизма копирования и вставки, или ТЕ класса II, которые могут быть вырезаны из генома и вставлены в новое место.
Движение ТЕ является движущей силой эволюции генома у эукариот, поскольку их вставка может нарушать функции генов, гомологичная рекомбинация между ТЕ может вызывать дупликации, а ТЕ может перемещать экзоны и регуляторные последовательности в новые места.[34]
Ретротранспозоны
Ретротранспозоны[35] обнаруживаются в основном у эукариот, но не встречаются у прокариот, а ретротранспозоны составляют значительную часть геномов многих эукариот. Ретротранспозон - это подвижный элемент, который переносится через РНК средний. Ретротранспозоны[36] состоят из ДНК, но транскрибируются в РНК для транспозиции, затем транскрипт РНК копируется обратно для образования ДНК с помощью определенного фермента, называемого обратной транскриптазой. Ретротранспозоны, которые несут обратную транскриптазу в своем гене, могут запускать собственную транспозицию, но гены, в которых отсутствует обратная транскриптаза, должны использовать обратную транскриптазу, синтезируемую другим ретротранспозоном. Ретротранспозоны могут быть транскрибированы в РНК, которые затем дублируются в другом сайте генома.[37] Ретротранспозоны можно разделить на длинные концевые повторы (LTR) и недлинные терминальные повторы (Non-LTR).[34]
Длинные концевые повторы (LTR) происходят от древних ретровирусных инфекций, поэтому они кодируют белки, связанные с ретровирусными белками, включая гены gag (структурные белки вируса), pol (обратная транскриптаза и интеграза), pro (протеаза) и в некоторых случаях env (оболочки).[33] Эти гены фланкированы длинными повторами как на 5 ', так и на 3' концах. Сообщалось, что LTR составляют наибольшую долю в геноме большинства растений и могут объяснять огромные различия в размере генома.[38]
Недлинные концевые повторы (Non-LTR) классифицируются как долго вкрапленные ядерные элементы (ЛИНИИ), короткие вкрапленные ядерные элементы (SINE) и элементы, подобные Пенелопе (PLE). В Dictyostelium discoideum, есть еще DIRS-подобные элементы, принадлежащие Non-LTR. Не-LTR широко распространены в геномах эукариот.[39]
Длинные вкрапленные элементы (LINE) кодируют гены обратной транскриптазы и эндонуклеазы, что делает их автономными мобильными элементами. В геноме человека около 500 000 СТРОК, что составляет около 17% генома.[40]
Короткие вкрапленные элементы (SINE) обычно состоят из менее 500 пар оснований и не автономны, поэтому для транспозиции они полагаются на белки, кодируемые LINE.[41] В Элемент Alu является наиболее распространенным синусоидальным синусом у приматов. Он составляет около 350 пар оснований и занимает около 11% генома человека с примерно 1 500 000 копий.[34]
Транспозоны ДНК
Транспозоны ДНК кодируют фермент транспозазу между инвертированными концевыми повторами. При экспрессии транспозаза распознает концевые инвертированные повторы, фланкирующие транспозон, и катализирует его вырезание и повторное внедрение в новый сайт.[30] Этот механизм вырезания и вставки обычно повторно вставляет транспозоны рядом с их исходным местоположением (в пределах 100 КБ).[34] Транспозоны ДНК находятся в бактериях и составляют 3% генома человека и 12% генома аскариды. C. elegans.[34]
Размер генома
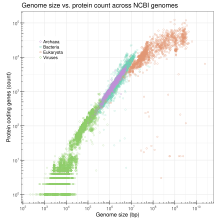
Размер генома - общее количество пар оснований ДНК в одной копии гаплоидного генома. Размер генома широко варьируется в зависимости от вида. У человека ядерный геном состоит примерно из 3,2 миллиарда нуклеотидов ДНК, разделенных на 24 линейных молекулы, самые короткие 50 000 000 нуклеотидов в длину и самые длинные 260 000 000 нуклеотидов, каждая из которых содержится в разных хромосомах.[42] Нет четкой и последовательной корреляции между морфологической сложностью и размером генома ни в одном из них. прокариоты или ниже эукариоты.[28][43] Размер генома в значительной степени является функцией расширения и сжатия повторяющихся элементов ДНК.
Поскольку геномы очень сложны, одна из исследовательских стратегий состоит в том, чтобы сократить количество генов в геноме до минимума и при этом сохранить жизнеспособность рассматриваемого организма. В настоящее время проводятся экспериментальные работы по минимальным геномам одноклеточных организмов, а также минимальным геномам для многоклеточных организмов (см. Биология развития ). Работа как in vivo и in silico.[44][45]
Вот таблица некоторых значимых или репрезентативных геномов. Видеть #Смотрите также для списков секвенированных геномов.
| Тип организма | Организм | Размер генома (пар оснований ) | Прибл. нет. генов | Примечание | |
|---|---|---|---|---|---|
| Вирус | Цирковирус свиней Тип 1 | 1,759 | 1,8 КБ | Самые маленькие вирусы, автономно реплицирующиеся в эукариотический клетки.[46] | |
| Вирус | Бактериофаг MS2 | 3,569 | 3,5 КБ | Первый секвенированный РНК-геном[47] | |
| Вирус | SV40 | 5,224 | 5,2 КБ | [48] | |
| Вирус | Фаг Φ-X174 | 5,386 | 5,4 КБ | Первый секвенированный ДНК-геном[49] | |
| Вирус | ВИЧ | 9,749 | 9,7 КБ | [50] | |
| Вирус | Фаг λ | 48,502 | 48,5 КБ | Часто используется как вектор для клонирования рекомбинантной ДНК. | |
| Вирус | Мегавирус | 1,259,197 | 1.3 Мб | До 2013 года самый крупный известный вирусный геном.[54] | |
| Вирус | Pandoravirus salinus | 2,470,000 | 2.47Мб | Самый большой известный вирусный геном.[55] | |
| Эукариотический органелла | Человек митохондрия | 16,569 | 16.6кб | [56] | |
| Бактерия | Nasuia deltocephalinicola (штамм NAS-ALF) | 112,091 | 112кб | 137 | Наименьший известный невирусный геном. Симбионт цикадки.[57] |
| Бактерия | Карсонелла руддии | 159,662 | 160кб | An эндосимбионт из псиллид насекомые | |
| Бактерия | Buchnera aphidicola | 600,000 | 600кб | Эндосимбионт тля[58] | |
| Бактерия | Вигглсуортия глоссинидия | 700,000 | 700 КБ | Симбионт в кишечнике муха цеце | |
| Бактерия – цианобактерии | Прохлорококк виды (1,7 Мб) | 1,700,000 | 1,7 Мб | 1,884 | Наименьший из известных геномов цианобактерий. Один из основных фотосинтезаторов на Земле.[59][60] |
| Бактерия | Haemophilus influenzae | 1,830,000 | 1.8 Мб | Секвенирование первого генома живого организма, июль 1995 г.[61] | |
| Бактерия | кишечная палочка | 4,600,000 | 4.6Мб | 4,288 | [62] |
| Бактерия - цианобактерии | Nostoc punctiforme | 9,000,000 | 9Мб | 7,432 | 7432 открытые рамки для чтения[63] |
| Бактерия | Solibacter usitatus (штамм Ellin 6076) | 9,970,000 | 10Мб | [64] | |
| Амебовидный | Полихаос дубиум ("Амеба" дубия) | 670,000,000,000 | 670Гб | Самый большой известный геном.[65] (Оспаривается)[66] | |
| Растение | Genlisea tuberosa | 61,000,000 | 61Мб | Наименьший зарегистрированный цветущее растение геном, 2014.[67] | |
| Растение | Arabidopsis thaliana | 135,000,000[68] | 135 Мб | 27,655[69] | Секвенирование первого генома растения, декабрь 2000 г.[70] |
| Растение | Populus trichocarpa | 480,000,000 | 480 Мб | 73,013 | Секвенирование первого генома дерева, сентябрь 2006 г.[71] |
| Растение | Fritillaria assyriaca | 130,000,000,000 | 130 ГБ | ||
| Растение | Париж японская (Родом из Японии, бледно-лепестковый) | 150,000,000,000 | 150 ГБ | Известен крупнейший геном растения[72] | |
| Растение – мох | Physcomitrella patens | 480,000,000 | 480 Мб | Первый геном мохообразный очередность, январь 2008 г.[73] | |
| Грибок – дрожжи | Saccharomyces cerevisiae | 12,100,000 | 12.1 Мб | 6,294 | Секвенирование первого эукариотического генома, 1996 г.[74] |
| Грибок | Aspergillus nidulans | 30,000,000 | 30Мб | 9,541 | [75] |
| Нематода | Pratylenchus coffeae | 20,000,000 | 20Мб | [76] Самый маленький известный геном животного[77] | |
| Нематода | Caenorhabditis elegans | 100,300,000 | 100Мб | 19,000 | Секвенирование первого генома многоклеточного животного, декабрь 1998 г.[78] |
| Насекомое | Drosophila melanogaster (плодовая муха) | 175,000,000 | 175 Мб | 13,600 | Размер варьируется в зависимости от деформации (175-180 МБ; стандартный y w напряжение 175Мб)[79] |
| Насекомое | Apis mellifera (пчела) | 236,000,000 | 236Мб | 10,157 | [80] |
| Насекомое | Bombyx mori (шелковая моль) | 432,000,000 | 432Мб | 14,623 | 14 623 предсказанных гена[81] |
| Насекомое | Solenopsis invicta (огненный муравей) | 480,000,000 | 480 Мб | 16,569 | [82] |
| Млекопитающее | Mus musculus | 2,700,000,000 | 2,7 ГБ | 20,210 | [83] |
| Млекопитающее | Пан панискус | 3,286,640,000 | 3,3 ГБ | 20,000 | Бонобо - предполагаемый размер генома 3,29 млрд п.н.[84] |
| Млекопитающее | Homo sapiens | 3,000,000,000 | 3Гб | 20,000 | Homo sapiens размер генома оценивается в 3,2 Гбит в 2001 г.[85][86] Начальная последовательность и анализ человеческого генома[87] |
| Птица | Gallus gallus | 1,043,000,000 | 1.0 ГБ | 20,000 | [88] |
| Рыбы | Тетраодон нигровиридис (вид рыбы фугу) | 385,000,000 | 390Мб | Наименьший известный геном позвоночных составляет 340 МБ.[89][90] - 385 Мб.[91] | |
| Рыбы | Протоптер эфиопический (мраморная двоякодышащая рыба) | 130,000,000,000 | 130 ГБ | Известен крупнейший геном позвоночных | |
Геномные изменения
Все клетки организма происходят из одной клетки, поэтому ожидается, что они будут иметь идентичные геномы; однако в некоторых случаях возникают различия. Как процесс копирования ДНК во время деления клеток, так и воздействие мутагенов окружающей среды могут привести к мутациям в соматических клетках. В некоторых случаях такие мутации приводят к раку, потому что они заставляют клетки быстрее делиться и вторгаться в окружающие ткани.[92] В некоторых лимфоцитах иммунной системы человека V (D) J рекомбинация генерирует различные геномные последовательности, так что каждая клетка продуцирует уникальное антитело или рецепторы Т-клеток.
В течение мейоз диплоидные клетки дважды делятся с образованием гаплоидных зародышевых клеток. Во время этого процесса рекомбинация приводит к перетасовке генетического материала из гомологичных хромосом, поэтому каждая гамета имеет уникальный геном.
Полногеномное перепрограммирование
Полногеномное перепрограммирование у мыши первичные половые клетки вовлекает эпигенетический стирание отпечатка, ведущее к тотипотентность. Перепрограммированию способствует активная Деметилирование ДНК, процесс, в котором ДНК базовая эксцизионная пластика путь.[93] Этот путь используется для стирания CpG метилирование (5mC) в первичных половых клетках. Стирание 5mC происходит через его преобразование в 5-гидроксиметилцитозин (5hmC) обусловлено высоким уровнем энзимов десять-одиннадцать диоксигеназ TET1 и TET2.[94]
Эволюция генома
Геномы - это больше, чем сумма гены и иметь черты характера, которые могут быть измеренный и изучены без ссылки на детали каких-либо конкретных генов и их продуктов. Исследователи сравнивают такие черты, как кариотип (номер хромосомы), размер генома, порядок генов, систематическая ошибка использования кодонов, и GC-контент чтобы определить, какие механизмы могли создать огромное разнообразие геномов, существующих сегодня (последние обзоры см. в Brown 2002; Saccone and Pesole 2003; Benfey and Protopapas 2004; Gibson and Muse 2004; Reese 2004; Gregory 2005).
Дубликаты играют важную роль в формировании генома. Дублирование может варьироваться от расширения короткие тандемные повторы, к дублированию кластера генов, и вплоть до дупликации целых хромосом или даже целые геномы. Такие дупликации, вероятно, фундаментальны для создания генетической новизны.
Горизонтальный перенос генов используется для объяснения того, почему часто существует крайнее сходство между небольшими частями геномов двух организмов, которые в остальном очень отдаленно связаны. Горизонтальный перенос генов, кажется, распространен среди многих микробы. Также, эукариотические клетки похоже, перенесли некоторый генетический материал из своих хлоропласт и митохондриальный геномы к их ядерным хромосомам. Последние эмпирические данные показывают, что вирусы и субвирусные РНК-сети играют важную роль в создании генетической новизны и естественного редактирования генома.
В художественной литературе
Произведения научной фантастики демонстрируют опасения по поводу доступности последовательностей генома.
Роман Майкла Крайтона 1990 года парк Юрского периода и последующие фильм расскажите историю миллиардера, который создает тематический парк клонированных динозавров на отдаленном острове, что привело к катастрофическим последствиям. Генетик извлекает ДНК динозавров из крови древних комаров и заполняет пробелы ДНК современных видов, чтобы создать несколько видов динозавров. Теоретика хаоса просят высказать свое экспертное мнение о безопасности создания экосистемы с динозаврами, и он неоднократно предупреждает, что результаты проекта будут непредсказуемыми и, в конечном итоге, неконтролируемыми. Эти предупреждения об опасностях использования геномной информации - основная тема книги.
Фильм 1997 года Гаттака действие происходит в футуристическом обществе, где геномы детей спроектированы таким образом, чтобы содержать наиболее идеальную комбинацию черт их родителей, а такие показатели, как риск сердечных заболеваний и прогнозируемая продолжительность жизни, документируются для каждого человека на основе его генома. Люди, рожденные вне программы евгеники, известные как «In-Valids», страдают от дискриминации и переходят к черным профессиям. Главный герой фильма - Недействительный, который пытается преодолеть предполагаемые генетические разногласия и осуществить свою мечту о работе космическим навигатором. Фильм предостерегает от будущего, в котором геномная информация питает предрассудки и крайние классовые различия между теми, кто может и не может позволить себе генетически модифицированных детей.[95]
Смотрите также
- Размер бактериального генома
- Криоконсервация генетических ресурсов животных
- Браузер генома
- Компилятор генома
- Топология генома
- Полногеномное исследование ассоциации
- Список секвенированных геномов животных
- Список секвенированных геномов архей
- Список секвенированных бактериальных геномов
- Список секвенированных эукариотических геномов
- Список секвенированных геномов грибов
- Список секвенированных геномов растений
- Список секвенированных пластомов
- Список секвенированных геномов протистов
- Метагеномика
- Микробиом
- Молекулярная эпидемиология
- Молекулярная патологическая эпидемиология
- Молекулярная патология
- Последовательность нуклеиновой кислоты
- Пангеном
- Точная медицина
- Регуляторный ген
- Sequenceome
- Секвенирование всего генома
Рекомендации
- ^ Брозиус, J (2009), «Фрагментированный ген», Летопись Нью-Йоркской академии наук, 1178 (1): 186–93, Bibcode:2009НЯСА1178..186Б, Дои:10.1111 / j.1749-6632.2009.05004.x, PMID 19845638, S2CID 8279434
- ^ Ридли М (2006). Геном: автобиография вида в 23 главах (PDF). Нью-Йорк: Многолетний Харпер. ISBN 978-0-06-019497-0. Архивировано из оригинал (PDF) 24 октября 2018 г.. Получено 11 мая 2016.
- ^ Винклер HL (1920). Verbreitung und Ursache der Parthenogenesis im Pflanzen- und Tierreiche. Йена: Верлаг Фишер.
- ^ "определение генома в Оксфордском словаре". Получено 25 марта 2014.
- ^ Ледерберг Дж., МакКрей А.Т. (2001). "'Ome Sweet 'Omics - Генеалогическая сокровищница слов " (PDF). Ученый. 15 (7). Архивировано из оригинал (PDF) 29 сентября 2006 г.
- ^ "Все о генах". www.beowulf.org.uk.
- ^ "Геномный дом". 8 декабря 2010 г.. Получено 27 января 2011.
- ^ Циммер С (18 декабря 2013 г.). «Окаменелость пальца ноги обеспечивает полный геном неандертальца». Нью-Йорк Таймс. Получено 18 декабря 2013.
- ^ Прюфер К., Расимо Ф., Паттерсон Н., Джей Ф., Санкарараман С., Сойер С. и др. (Январь 2014). «Полная последовательность генома неандертальца с Горного Алтая». Природа. 505 (7481): 43–49. Bibcode:2014Натура.505 ... 43П. Дои:10.1038 / природа12886. ЧВК 4031459. PMID 24352235.
- ^ Уэйд Н. (31 мая 2007 г.). «Расшифрован геном первопроходца ДНК». Нью-Йорк Таймс. Получено 2 апреля 2010.
- ^ "Что такое геном?". Genomenewsnetwork.org. 15 января 2003 г.. Получено 27 января 2011.
- ^ NCBI_user_services (29 марта 2004 г.). «Картографический информационный бюллетень». Архивировано из оригинал 19 июля 2010 г.. Получено 27 января 2011.
- ^ Консорциум ссылок на геном. «Сборка генома». Получено 23 августа 2016.
- ^ Каплан, Сара (17 апреля 2016 г.). «Как ваши 20 000 генов определяют столько совершенно разных черт? Они многозадачны». Вашингтон Пост. Получено 27 августа 2016.
- ^ Отметьте Хайден, Эрика (2015). "Ученые надеются привлечь миллионы людей к" DNA.LAND "'". Природа. Дои:10.1038 / природа.2015.18514.
- ^ Циммер, Карл. «Игра геномов, серия 13: ответы и вопросы». СТАТ. Получено 27 августа 2016.
- ^ Гелдерблом, Ханс Р. (1996). Медицинская микробиология (4-е изд.). Галвестон, Техас: Медицинский филиал Техасского университета в Галвестоне.
- ^ Урри, Лиза А. (2016). Кэмпбелл Биология: седьмое издание. Нью-Йорк: Хобокен: Высшее образование Пирсона. С. 403–404. ISBN 0134093410.
- ^ Урри, Лиза А (2016). Кэмпбелл Биология: седьмое издание. Нью-Йорк: Хобокен: Высшее образование Пирсона. С. 403–404. ISBN 0134093410.
- ^ Самсон Р. Я., Белл С. Д. (2014). «Биология архейных хромосом». Журнал молекулярной микробиологии и биотехнологии. 24 (5–6): 420–27. Дои:10.1159/000368854. ЧВК 5175462. PMID 25732343.
- ^ Чаконас Дж., Чен К.В. (2005). "Репликация линейных бактериальных хромосом: больше не ходить по кругу". Бактериальная хромосома: 525–540. Дои:10.1128 / 9781555817640.ch29. ISBN 9781555812324.
- ^ «Бактериальные хромосомы». Микробная генетика. 2002.
- ^ а б Кунин Е.В., Вольф Ю.И. (июль 2010 г.). «Ограничения и пластичность в эволюции генома и молекулярного феномена». Обзоры природы. Генетика. 11 (7): 487–98. Дои:10.1038 / nrg2810. ЧВК 3273317. PMID 20548290.
- ^ McCutcheon JP, Moran NA (ноябрь 2011 г.). «Экстремальное сокращение генома симбиотических бактерий». Обзоры природы. Микробиология. 10 (1): 13–26. Дои:10.1038 / nrmicro2670. PMID 22064560. S2CID 7175976.
- ^ Land M, Hauser L, Jun SR, Nookaew I, Leuze MR, Ahn TH, Karpinets T, Lund O, Kora G, Wassenaar T, Poudel S, Ussery DW (март 2015 г.). «Выводы за 20 лет секвенирования бактериального генома». Функциональная и интегративная геномика. 15 (2): 141–61. Дои:10.1007 / s10142-015-0433-4. ЧВК 4361730. PMID 25722247.
- ^ "Ученые обнаружили бесполого крошечного червяка, чья родословная насчитывает 18 миллионов лет". ScienceDaily. Получено 7 ноября 2017.
- ^ Khandelwal S (март 1990 г.). «Хромосомная эволюция рода Ophioglossum L.». Ботанический журнал Линнеевского общества. 102 (3): 205–17. Дои:10.1111 / j.1095-8339.1990.tb01876.x.
- ^ а б c Левин Б. (2004). Гены VIII (8-е изд.). Река Аппер Сэдл, Нью-Джерси: Пирсон / Прентис-Холл. ISBN 978-0-13-143981-8.
- ^ Стоянович Н, изд. (2007). Вычислительная геномика: современные методы. Wymondham: Horizon Bioscience. ISBN 978-1-904933-30-4.
- ^ а б c Падекен Дж., Целлер П., Гассер С.М. (апрель 2015 г.). «Повторите ДНК в организации и стабильности генома». Текущее мнение в области генетики и развития. 31: 12–19. Дои:10.1016 / j.gde.2015.03.009. PMID 25917896.
- ^ а б Усдин К. (июль 2008 г.). «Биологические эффекты простых тандемных повторов: уроки болезней распространения повторений». Геномные исследования. 18 (7): 1011–19. Дои:10.1101 / гр.070409.107. ЧВК 3960014. PMID 18593815.
- ^ Ли Ю.К., Король А.Б., Фахима Т., Бейлес А., Нево Е. (декабрь 2002 г.). «Микросателлиты: геномное распределение, предполагаемые функции и мутационные механизмы: обзор». Молекулярная экология. 11 (12): 2453–65. Дои:10.1046 / j.1365-294X.2002.01643.x. PMID 12453231. S2CID 23606208.
- ^ а б Wessler SR (ноябрь 2006 г.). «Мобильные элементы и эволюция геномов эукариот». Труды Национальной академии наук Соединенных Штатов Америки. 103 (47): 17600–01. Bibcode:2006PNAS..10317600W. Дои:10.1073 / pnas.0607612103. ЧВК 1693792. PMID 17101965.
- ^ а б c d е Казазян HH (март 2004 г.). «Мобильные элементы: драйверы эволюции генома». Наука. 303 (5664): 1626–32. Bibcode:2004Наука ... 303.1626K. Дои:10.1126 / science.1089670. PMID 15016989. S2CID 1956932.
- ^ "Транспозон | генетика". Энциклопедия Британника. Получено 5 декабря 2020.
- ^ Сандерс, Марк Фредерик (2019). Генетический анализ: комплексный подход третье издание. Нью-Йорк: Пирсон, всегда учится и совершенствуется. п. 425. ISBN 9780134605173.
- ^ Deininger PL, Moran JV, Batzer MA, Kazazian HH (декабрь 2003 г.). «Мобильные элементы и эволюция генома млекопитающих». Текущее мнение в области генетики и развития. 13 (6): 651–58. Дои:10.1016 / j.gde.2003.10.013. PMID 14638329.
- ^ Кидвелл М.Г., Лиш Д.Р. (март 2000 г.). «Мобильные элементы и эволюция генома хозяина». Тенденции в экологии и эволюции. 15 (3): 95–99. Дои:10.1016 / S0169-5347 (99) 01817-0. PMID 10675923.
- ^ Ричард Г.Ф., Керрест А., Дуйон Б. (декабрь 2008 г.). «Сравнительная геномика и молекулярная динамика повторов ДНК у эукариот». Обзоры микробиологии и молекулярной биологии. 72 (4): 686–727. Дои:10.1128 / MMBR.00011-08. ЧВК 2593564. PMID 19052325.
- ^ Cordaux R, Batzer MA (октябрь 2009 г.). «Влияние ретротранспозонов на эволюцию генома человека». Обзоры природы. Генетика. 10 (10): 691–703. Дои:10.1038 / nrg2640. ЧВК 2884099. PMID 19763152.
- ^ Хан Дж. С., Боке Дж. Д. (август 2005 г.). «Ретротранспозоны LINE-1: модуляторы количества и качества экспрессии генов млекопитающих?». BioEssays. 27 (8): 775–84. Дои:10.1002 / bies.20257. PMID 16015595. S2CID 26424042.
- ^ "Человеческий геном". Получено 19 августа 2016.
- ^ Грегори Т.Р., Никол Дж.А., Тамм Х., Куллман Б., Куллман К., Лейтч И.Дж., Мюррей Б.Г., Капраун Д.Ф., Грейлхубер Дж., Беннетт, доктор медицины (январь 2007 г.). «Базы данных размеров эукариотических геномов». Исследования нуклеиновых кислот. 35 (Выпуск базы данных): D332–38. Дои:10.1093 / нар / gkl828. ЧВК 1669731. PMID 17090588.
- ^ Гласс Д.И., Асад-Гарсия Н., Альперович Н., Юсеф С., Льюис М.Р., Маруф М., Хатчисон, Калифорния, Смит Х.о., Вентер Дж.С. (январь 2006 г.). «Основные гены минимальной бактерии». Труды Национальной академии наук Соединенных Штатов Америки. 103 (2): 425–30. Bibcode:2006ПНАС..103..425Г. Дои:10.1073 / pnas.0510013103. ЧВК 1324956. PMID 16407165.
- ^ Форстер AC, Church GM (2006). «К синтезу минимальной клетки». Молекулярная системная биология. 2 (1): 45. Дои:10.1038 / msb4100090. ЧВК 1681520. PMID 16924266.
- ^ Манкерц П. (2008). «Молекулярная биология цирковирусов свиней». Вирусы животных: молекулярная биология. Caister Academic Press. ISBN 978-1-904455-22-6.
- ^ Фирс В., Контрерас Р., Дуеринк Ф., Хегеман Г., Изерентант Д., Меррегерт Дж., Мин Джоу В., Молеманс Ф., Рэймакерс А., Ван ден Берге А., Фолькаерт Г., Исебаерт М. (апрель 1976 г.). «Полная нуклеотидная последовательность РНК бактериофага MS2: первичная и вторичная структура гена репликазы». Природа. 260 (5551): 500–07. Bibcode:1976Натура.260..500F. Дои:10.1038 / 260500a0. PMID 1264203. S2CID 4289674.
- ^ Fiers W, Contreras R, Haegemann G, Rogiers R, Van de Voorde A, Van Heuverswyn H, Van Herreweghe J, Volckaert G, Ysebaert M (май 1978). «Полная нуклеотидная последовательность ДНК SV40». Природа. 273 (5658): 113–20. Bibcode:1978Натура.273..113F. Дои:10.1038 / 273113a0. PMID 205802. S2CID 1634424.
- ^ Сэнгер Ф., Air GM, Баррелл Б.Г., Браун Н.Л., Колсон А.Р., Фиддес, Калифорния, Хатчисон, Калифорния, Слокомб П.М., Смит М. (февраль 1977 г.). «Нуклеотидная последовательность ДНК бактериофага phi X174». Природа. 265 (5596): 687–95. Bibcode:1977Натура.265..687С. Дои:10.1038 / 265687a0. PMID 870828. S2CID 4206886.
- ^ «Вирусология - вирус иммунодефицита человека и СПИД, структура: геном и белки ВИЧ». Pathmicro.med.sc.edu. 1 июля 2010 г.. Получено 27 января 2011.
- ^ Томасон Л., Суд Д.Л., Бубуненко М., Константино Н., Уилсон Х., Датта С., Оппенгейм А. (апрель 2007 г.). «Рекомбинирование: генная инженерия бактерий с использованием гомологичной рекомбинации». Текущие протоколы в молекулярной биологии. Глава 1: Раздел 1.16. Дои:10.1002 / 0471142727.mb0116s78. ISBN 978-0-471-14272-0. PMID 18265390. S2CID 490362.
- ^ Суд DL, Oppenheim AB, Adhya SL (январь 2007 г.). «Новый взгляд на генетические сети лямбда бактериофагов». Журнал бактериологии. 189 (2): 298–304. Дои:10.1128 / JB.01215-06. ЧВК 1797383. PMID 17085553.
- ^ Сангер Ф., Колсон А. Р., Хонг Г. Ф., Хилл Д. Ф., Петерсен ГБ (декабрь 1982 г.). «Нуклеотидная последовательность лямбда-ДНК бактериофага». Журнал молекулярной биологии. 162 (4): 729–73. Дои:10.1016/0022-2836(82)90546-0. PMID 6221115.
- ^ Лежандр М., Арслан Д., Абергель С., Клавери Дж. М. (январь 2012 г.). «Геномика мегавируса и неуловимая четвертая область жизни». Коммуникативная и интегративная биология. 5 (1): 102–06. Дои:10.4161 / cib.18624. ЧВК 3291303. PMID 22482024.
- ^ Филипп Н., Лежандр М., Дутре Дж., Кутэ Y, Пуаро О, Леско М., Арслан Д., Зельцер В., Берто Л., Брюли С., Гарин Дж., Клавери Дж. М., Абергель С. (июль 2013 г.). «Пандоровирусы: вирусы амеб с геномом до 2,5 Мб, достигающие генома паразитических эукариот» (PDF). Наука. 341 (6143): 281–86. Bibcode:2013Научный ... 341..281П. Дои:10.1126 / science.1239181. PMID 23869018. S2CID 16877147.
- ^ Андерсон С., Банкир А.Т., Баррелл Б.Г., де Брейн М.Х., Коулсон А.Р., Друин Дж., Эперон И.К., Нирлих Д.П., Роу Б.А., Сэнгер Ф., Шрайер П.Х., Смит А.Дж., Стаден Р., Янг И.Г. (апрель 1981 г.). «Последовательность и организация митохондриального генома человека». Природа. 290 (5806): 457–65. Bibcode:1981Натура.290..457A. Дои:10.1038 / 290457a0. PMID 7219534. S2CID 4355527.
- ^ Беннетт Г.М., Моран Н.А. (5 августа 2013 г.). «Маленький, меньший, самый маленький: происхождение и эволюция древних двойных симбиозов у насекомых, питающихся флоэмами». Геномная биология и эволюция. 5 (9): 1675–88. Дои:10.1093 / gbe / evt118. ЧВК 3787670. PMID 23918810.
- ^ Сигенобу С., Ватанабэ Х., Хаттори М., Сакаки Ю., Исикава Х. (сентябрь 2000 г.). «Последовательность генома внутриклеточного бактериального симбионта тлей Buchnera sp. APS». Природа. 407 (6800): 81–86. Bibcode:2000Натура 407 ... 81С. Дои:10.1038/35024074. PMID 10993077.
- ^ Rocap G, Larimer FW, Lamerdin J, Malfatti S, Chain P, Ahlgren NA и др. (Август 2003 г.). «Дивергенция генома у двух экотипов Prochlorococcus отражает дифференциацию океанических ниш». Природа. 424 (6952): 1042–47. Bibcode:2003Натура.424.1042R. Дои:10.1038 / природа01947. PMID 12917642. S2CID 4344597.
- ^ Dufresne A, Salanoubat M, Partensky F, Artiguenave F, Axmann IM, Barbe V и др. (Август 2003 г.). «Последовательность генома цианобактерии Prochlorococcus marinus SS120, почти минимальный оксифототрофный геном». Труды Национальной академии наук Соединенных Штатов Америки. 100 (17): 10020–25. Bibcode:2003ПНАС..10010020Д. Дои:10.1073 / pnas.1733211100. ЧВК 187748. PMID 12917486.
- ^ Флейшманн Р.Д., Адамс, доктор медицины, Уайт О., Клейтон Р.А., Киркнесс Э.Ф., Керлаваж А.Р., Булт С.Дж., Могила Д.Ф., Догерти Б.А., Меррик Дж.М. (июль 1995 г.). «Полногеномное случайное секвенирование и сборка Haemophilus influenzae Rd». Наука. 269 (5223): 496–512. Bibcode:1995Научный ... 269..496F. Дои:10.1126 / science.7542800. PMID 7542800. S2CID 10423613.
- ^ Blattner FR, Plunkett G, Bloch CA, Perna NT, Burland V, Riley M и др. (Сентябрь 1997 г.). «Полная последовательность генома Escherichia coli K-12». Наука. 277 (5331): 1453–62. Дои:10.1126 / science.277.5331.1453. PMID 9278503.
- ^ Микс Дж. К., Эльхай Дж., Тиль Т., Поттс М., Лаример Ф., Ламердин Дж., Предки П., Атлас Р. (2001). «Обзор генома Nostoc punctiforme, многоклеточной симбиотической цианобактерии». Фотосинтез Исследования. 70 (1): 85–106. Дои:10.1023 / А: 1013840025518. PMID 16228364. S2CID 8752382.
- ^ Challacombe JF, Eichorst SA, Hauser L, Land M, Xie G, Kuske CR (15 сентября 2011 г.). Стейнке Д. (ред.). «Биологические последствия приобретения и дупликации древних генов в большом геноме Candidatus Solibacter usitatus Ellin6076». PLOS ONE. 6 (9): e24882. Bibcode:2011PLoSO ... 624882C. Дои:10.1371 / journal.pone.0024882. ЧВК 3174227. PMID 21949776.
- ^ Парфри Л.В., Лар DJ, Кац Л.А. (апрель 2008 г.). «Динамическая природа геномов эукариот». Молекулярная биология и эволюция. 25 (4): 787–94. Дои:10.1093 / molbev / msn032. ЧВК 2933061. PMID 18258610.
- ^ ScienceShot: самый крупный геном в истории В архиве 11 октября 2010 г. Wayback Machine, комментирует: «Измерение Amoeba dubia и других простейших, которые, как сообщается, имеют очень большие геномы, было сделано в 1960-х годах с использованием грубого биохимического подхода, который сейчас считается ненадежным методом для точного определения размера генома».
- ^ Fleischmann A, Michael TP, Rivadavia F, Sousa A, Wang W, Temsch EM, Greilhuber J, Müller KF, Heubl G (декабрь 2014 г.). «Эволюция размера генома и числа хромосом у плотоядных растений рода Genlisea (Lentibulariaceae) с новой оценкой минимального размера генома у покрытосеменных». Анналы ботаники. 114 (8): 1651–63. Дои:10.1093 / aob / mcu189. ЧВК 4649684. PMID 25274549.
- ^ «Сборка генома». Информационный ресурс по арабидопсису (ТАИР).
- ^ «Детали - Arabidopsis thaliana - Ensembl Genomes 40». растения.ensembl.org.
- ^ Грейлхубер Дж., Борщ Т., Мюллер К., Ворберг А., Порембски С., Бартлотт В. (ноябрь 2006 г.). «Наименьшие геномы покрытосеменных, обнаруженные у lentibulariaceae, с хромосомами размером с бактерии». Биология растений. 8 (6): 770–77. Дои:10.1055 / с-2006-924101. PMID 17203433.
- ^ Тускан Г.А., Дифацио С., Янссон С., Больманн Дж., Григорьев И., Хелльстен Ю. и др. (Сентябрь 2006 г.). «Геном тополя черного, Populus trichocarpa (Torr. & Gray)» (PDF). Наука. 313 (5793): 1596–604. Bibcode:2006Научный ... 313.1596Т. Дои:10.1126 / science.1128691. PMID 16973872. S2CID 7717980.
- ^ Пеллисер Дж., Фэй М.Ф., Лейтч И.Дж. (15 сентября 2010 г.). "Самый крупный из них эукариотический геном?". Ботанический журнал Линнеевского общества. 164 (1): 10–15. Дои:10.1111 / j.1095-8339.2010.01072.x.
- ^ Ланг Д., Циммер А.Д., Ренсинг С.А., Рески Р. (октябрь 2008 г.). «Изучение биоразнообразия растений: геном Physcomitrella и не только». Тенденции в растениеводстве. 13 (10): 542–49. Дои:10.1016 / j.tplants.2008.07.002. PMID 18762443.
- ^ "База данных генома сахаромицетов". Yeastgenome.org. Получено 27 января 2011.
- ^ Галаган Дж. Э., Кальво С. Е., Куомо С., Ма Л. Дж., Вортман Дж. Р., Батцоглу С. и др. (Декабрь 2005 г.). «Секвенирование Aspergillus nidulans и сравнительный анализ с A. fumigatus и A. oryzae». Природа. 438 (7071): 1105–15. Bibcode:2005 Натур.438.1105Г. Дои:10.1038 / природа04341. PMID 16372000.
- ^ Лерой С., Буамер С., Моран С., Фаржетт М. (2007). «Размер генома нематод, паразитирующих на растениях». Нематология. 9 (3): 449–50. Дои:10.1163/156854107781352089.
- ^ Грегори Т.Р. (2005). «База данных размеров генома животных». Грегори, Т. (2016). База данных размеров генома животных.
- ^ В C. elegans Консорциум по секвенированию (декабрь 1998 г.). «Последовательность генома нематоды C. elegans: платформа для изучения биологии». Наука. 282 (5396): 2012–18. Bibcode:1998На ... 282.2012.. Дои:10.1126 / science.282.5396.2012. PMID 9851916. S2CID 16873716.
- ^ Эллис Л.Л., Хуанг В., Куинн А.М., Ахуджа А., Альфрейд Б., Гомес Ф.Э., Хьельмен С.Э., Мур К.Л., Маккей Т.Ф., Джонстон Дж.С., Тарон А.М. (июль 2014 г.). «Внутрипопуляционные вариации размера генома D. melanogaster отражают вариабельность и пластичность жизненного цикла». PLOS Genetics. 10 (7): e1004522. Дои:10.1371 / journal.pgen.1004522. ЧВК 4109859. PMID 25057905.
- ^ Консорциум по секвенированию генома медоносной пчелы (октябрь 2006 г.). «Понимание социальных насекомых из генома пчелы Apis mellifera». Природа. 443 (7114): 931–49. Bibcode:2006Натура.443..931Т. Дои:10.1038 / природа05260. ЧВК 2048586. PMID 17073008.
- ^ Международный геном шелкопряда (декабрь 2008 г.). «Геном чешуекрылого модельного насекомого, тутового шелкопряда Bombyx mori». Биохимия и молекулярная биология насекомых. 38 (12): 1036–45. Дои:10.1016 / j.ibmb.2008.11.004. PMID 19121390.
- ^ Wurm Y, Wang J, Riba-Grognuz O, Corona M, Nygaard S, Hunt BG и др. (Апрель 2011 г.). «Геном огненного муравья Solenopsis invicta». Труды Национальной академии наук Соединенных Штатов Америки. 108 (14): 5679–84. Bibcode:2011PNAS..108.5679W. Дои:10.1073 / pnas.1009690108. ЧВК 3078418. PMID 21282665.
- ^ Черч Д.М., Гудштадт Л., Хиллиер Л.В., Зоди М.С., Гольдштейн С., Ше Икс и др. (Май 2009 г.). Робертс Р.Дж. (ред.). «Специфическая биология линии, выявленная завершенной сборкой генома мыши». PLOS Биология. 7 (5): e1000112. Дои:10.1371 / journal.pbio.1000112. ЧВК 2680341. PMID 19468303.
- ^ «Пан панискус (карликовый шимпанзе)». nih.gov. Получено 30 июн 2016.
- ^ Эрик Ландер; и другие. (15 февраля 2001 г.). "Начальная последовательность и анализ человеческого генома". Природа. 409 (6822): 860–921. Дои:10.1038/35057062. PMID 11237011. Таблица 8.
- ^ «Информационный бюллетень по функциональной и сравнительной геномике». Ornl.gov. Архивировано из оригинал 20 сентября 2008 г.
- ^ Venter JC, Адамс, доктор медицины, Майерс Е.В., Ли П.В., Фреска Р.Дж., Саттон Г.Г. и др. (Февраль 2001 г.). «Последовательность генома человека». Наука. 291 (5507): 1304–51. Bibcode:2001Научный ... 291.1304V. Дои:10.1126 / science.1058040. PMID 11181995.
- ^ Международный консорциум по секвенированию куриного генома (декабрь 2004 г.). «Последовательность и сравнительный анализ генома курицы дают уникальные перспективы эволюции позвоночных». Природа. 432 (7018): 695–716. Bibcode:2004Натура 432..695С. Дои:10.1038 / природа03154. ISSN 0028-0836. PMID 15592404.
- ^ Руст Кроллиус Х, Джайон О., Дасильва С., Озуф-Костаз С., Физамес С., Фишер С., Буно Л., Бийо А., Кетье Ф, Саурин В., Бернот А., Вайссенбах Дж. (Июль 2000 г.). «Характеристика и повторный анализ компактного генома пресноводного иглобрюха Tetraodon nigroviridis». Геномные исследования. 10 (7): 939–49. Дои:10.1101 / гр. 10.7.939. ЧВК 310905. PMID 10899143.
- ^ Jaillon O, Aury JM, Brunet F, Petit JL, Stange-Thomann N, Mauceli E, et al. (Октябрь 2004 г.). «Дублирование генома костистых рыб Tetraodon nigroviridis выявляет ранний протокариотип позвоночных». Природа. 431 (7011): 946–57. Bibcode:2004 Натур.431..946J. Дои:10.1038 / природа03025. PMID 15496914.
- ^ "Информация о проекте Tetraodon". Архивировано из оригинал 26 сентября 2012 г.. Получено 17 октября 2012.
- ^ Мартинкорена I, Кэмпбелл П.Дж. (сентябрь 2015 г.). «Соматическая мутация в раковых и нормальных клетках». Наука. 349 (6255): 1483–89. Bibcode:2015Научный ... 349.1483M. Дои:10.1126 / science.aab4082. PMID 26404825. S2CID 13945473.
- ^ Хайкова П., Джеффрис С.Дж., Ли С., Миллер Н., Джексон С.П., Сурани М.А. (июль 2010 г.). «Полногеномное перепрограммирование в зародышевой линии мыши влечет за собой основной путь эксцизионной репарации». Наука. 329 (5987): 78–82. Bibcode:2010Sci ... 329 ... 78H. Дои:10.1126 / science.1187945. ЧВК 3863715. PMID 20595612.
- ^ Hackett JA, Sengupta R, Zylicz JJ, Murakami K, Lee C, Down TA, Surani MA (январь 2013 г.). «Динамика деметилирования ДНК зародышевой линии и стирание отпечатка с помощью 5-гидроксиметилцитозина». Наука. 339 (6118): 448–52. Bibcode:2013Наука ... 339..448H. Дои:10.1126 / science.1229277. ЧВК 3847602. PMID 23223451.
- ^ «Гаттака (фильм)». Гнилые помидоры.
дальнейшее чтение
- Бенфей П., Протопапас А.Д. (2004). Основы геномики. Прентис Холл.
- Браун Т.А. (2002). Геномы 2. Оксфорд: Bios Scientific Publishers. ISBN 978-1-85996-029-5.
- Гибсон Дж., Муза С.В. (2004). Учебник по геномной науке (Второе изд.). Сандерленд, Массачусетс: Sinauer Assoc. ISBN 978-0-87893-234-4.
- Грегори Т.Р. (2005). Эволюция генома. Эльзевир. ISBN 978-0-12-301463-4.
- Рис RJ (2004). Analysis of Genes and Genomes. Чичестер: Джон Уайли и сыновья. ISBN 978-0-470-84379-6.
- Saccone C, Pesole G (2003). Handbook of Comparative Genomics. Чичестер: Джон Уайли и сыновья. ISBN 978-0-471-39128-9.
- Werner E (December 2003). "In silico multicellular systems biology and minimal genomes". Открытие наркотиков сегодня. 8 (24): 1121–27. Дои:10.1016/S1359-6446(03)02918-0. PMID 14678738.
внешняя ссылка
- Браузер генома UCSC – view the genome and annotations for more than 80 organisms.
- genomecenter.howard.edu
- Build a DNA Molecule
- Some comparative genome sizes
- DNA Interactive: The History of DNA Science
- DNA From The Beginning
- All About The Human Genome Project —from Genome.gov
- Animal genome size database
- Plant genome size database
- GOLD:Genomes OnLine Database
- The Genome News Network
- NCBI Entrez Genome Project database
- NCBI Genome Primer
- Генные Карты —an integrated database of human genes
- BBC News – Final genome 'chapter' published
- IMG (The Integrated Microbial Genomes system)—for genome analysis by the DOE-JGI
- GeKnome Technologies Next-Gen Sequencing Data Analysis —next-generation sequencing data analysis for Иллюмина и 454 Service from GeKnome Technologies.
| Ключевые компоненты | |
|---|---|
| Поля | |
| Археогенетика из | |
| похожие темы | |
| Списки | |
| |
| Genomics | |
|---|---|
| Биоинформатика | |
| Структурная биология | |
| Инструменты исследования | |
| Организации |
|
| |