Протеин - Protein

Белки большие биомолекулы, или же макромолекулы, состоящий из одной или нескольких длинных цепочек аминокислота остатки. Белки выполняют в организмах широкий спектр функций, в том числе катализатор метаболических реакций, Репликация ДНК, реагируя на раздражители, предоставляя структура к ячейкам и организмы, и транспортирующие молекулы из одного места в другое. Белки отличаются друг от друга прежде всего последовательностью аминокислот, которая продиктована нуклеотидная последовательность от их гены, что обычно приводит к сворачивание белка в конкретный 3D структура что определяет его деятельность.
Линейная цепочка аминокислотных остатков называется полипептид. Белок содержит по крайней мере один длинный полипептид. Короткие полипептиды, содержащие менее 20–30 остатков, редко считаются белками и обычно называются пептиды, а иногда олигопептиды. Отдельные аминокислотные остатки связаны вместе пептидные связи и соседние аминокислотные остатки. В последовательность аминокислотных остатков в белке определяется последовательность из ген, который закодирован в генетический код. В общем, генетический код определяет 20 стандартных аминокислот; но у некоторых организмов генетический код может включать селеноцистеин и - в некоторых археи —пирролизин. Вскоре после или даже во время синтеза остатки в белке часто химически модифицируются с помощью посттрансляционная модификация, который изменяет физические и химические свойства, укладку, стабильность, активность и, в конечном итоге, функцию белков. К некоторым белкам присоединены непептидные группы, которые можно назвать протезные группы или же кофакторы. Белки также могут работать вместе для достижения определенной функции, и они часто объединяются, образуя стабильные белковые комплексы.
После образования белки существуют только в течение определенного периода, а затем деградированный и перерабатывается механизмами клетки в процессе белковый оборот. Продолжительность жизни протеина измеряется его период полураспада и охватывает широкий спектр. Они могут существовать в течение минут или лет со средней продолжительностью жизни 1-2 дня в клетках млекопитающих. Аномальные или неправильно свернутые белки разлагаются быстрее либо из-за того, что они нацелены на разрушение, либо из-за нестабильности.
Как и другие биологические макромолекулы, такие как полисахариды и нуклеиновые кислоты, белки являются важными частями организмов и участвуют практически в каждом процессе внутри клетки. Многие белки ферменты который катализировать биохимические реакции и жизненно важны для метаболизм. Белки также выполняют структурные или механические функции, такие как актин и миозин в мышцах и белках в цитоскелет, которые образуют систему строительные леса который поддерживает форму клеток. Другие белки важны для клеточная сигнализация, иммунные ответы, клеточная адгезия, а клеточный цикл. У животных белки необходимы в рацион питания предоставить незаменимые аминокислоты этого не может быть синтезированный. Пищеварение расщепляет белки для использования в метаболизме.
Белки могут быть очищенный из других клеточных компонентов с использованием различных методов, таких как ультрацентрифугирование, осадки, электрофорез, и хроматография; появление генная инженерия сделал возможным ряд методов для облегчения очистки. Методы, обычно используемые для изучения структуры и функции белков, включают: иммуногистохимия, сайт-направленный мутагенез, Рентгеновская кристаллография, ядерный магнитный резонанс и масс-спектрометрии.
История и этимология
Белки были признаны отдельным классом биологических молекул в восемнадцатом веке. Антуан Фуркрой и другие, отличающиеся способностью молекул к коагулировать или же флокулировать под воздействием тепла или кислоты.[1] Известные примеры в то время включали альбумин из яичные белки, кровь сывороточный альбумин, фибрин, и пшеница глютен.
Белки были впервые описаны голландским химиком. Герардус Йоханнес Малдер и назван шведским химиком Йенс Якоб Берцелиус в 1838 г.[2][3] Малдер выполнил элементный анализ общих белков и обнаружил, что почти все белки имеют одинаковые эмпирическая формула, С400ЧАС620N100О120п1S1.[4] Он пришел к ошибочному выводу, что они могут состоять из одного типа (очень больших) молекул. Термин «белок» для описания этих молекул был предложен сотрудником Малдера Берцелиусом; белок получен из Греческий слово πρώτειος (Proteios), что означает "первичный",[5] "впереди" или "впереди",[6] + -в. Малдер продолжил идентифицировать продукты распада белка, такие как аминокислота лейцин для которого он нашел (почти правильный) молекулярный вес 131 Да.[4] До слова «белок» использовались другие названия, например, «альбумины» или «белковые материалы» (Eiweisskörper, на немецком).[7]
Ранние ученые-диетологи, такие как немец Карл фон Войт считали, что белок является самым важным питательным веществом для поддержания структуры тела, потому что обычно считалось, что «плоть рождает плоть».[8] Карл Генрих Риттхаузен расширенные известные формы белка с идентификацией глютаминовая кислота. На Сельскохозяйственная экспериментальная станция Коннектикута подробный обзор растительных белков был составлен Томас Бёрр Осборн. Работаю с Лафайет Мендель и применяя Закон минимума Либиха в кормлении лабораторные крысы, питательно незаменимые аминокислоты были созданы. Работа была продолжена и доведена до сведения Уильям Камминг Роуз. Понимание белков как полипептиды пришел через работу Франц Хофмайстер и Герман Эмиль Фишер в 1902 г.[9][10] Центральная роль белков как ферменты в живых организмах не была полностью оценена до 1926 г., когда Джеймс Б. Самнер показали, что фермент уреаза на самом деле был протеином.[11]
Из-за сложности очистки белков в больших количествах их изучение было очень трудным для ранних биохимиков. Следовательно, ранние исследования были сосредоточены на белках, которые можно было очищать в больших количествах, например, белков крови, яичного белка, различных токсинов и пищеварительных / метаболических ферментов, полученных на бойнях. В 1950-х годах Armor Hot Dog Co. очищенный 1 кг чистого бычьего панкреатического рибонуклеаза А и сделал его бесплатным для ученых; этот жест помог рибонуклеазе А стать главной целью биохимических исследований на следующие десятилетия.[4]

Линус Полинг приписывают успешное предсказание регулярного белка второстепенные конструкции на основе водородная связь, идея, впервые выдвинутая Уильям Эстбери в 1933 г.[12] Позже работа Вальтер Каузманн на денатурация,[13][14] частично на основе предыдущих исследований Кай Линдерстрём-Ланг,[15] способствовал пониманию сворачивание белка и структура опосредована гидрофобные взаимодействия.
Первый протеин, который будет последовательный был инсулин, к Фредерик Сэнгер, в 1949 году. Сэнгер правильно определил аминокислотную последовательность инсулина, убедительно продемонстрировав, что белки состоят из линейных полимеров аминокислот, а не из разветвленных цепей, коллоиды, или же циклолы.[16] За это достижение он получил Нобелевскую премию в 1958 году.[17]
Первый белковые структуры быть решенными были гемоглобин и миоглобин, к Макс Перуц и Сэр Джон Каудери Кендрю соответственно, в 1958 г.[18][19] По состоянию на 2017 год[Обновить], то Банк данных белков имеет более 126 060 структур белков с атомным разрешением.[20] В последнее время криоэлектронная микроскопия большого макромолекулярные сборки[21] и вычислительные предсказание структуры белка небольшого белка домены[22] два метода, приближающихся к атомарному разрешению.
Количество белков, закодированных в геномах
Количество белков, закодированных в геном примерно соответствует количеству гены (хотя может быть значительное количество генов, кодирующих РНК белка, например рибосомная РНК с). Вирусы обычно кодируют от нескольких до нескольких сотен белков, археи и бактерии от нескольких сотен до нескольких тысяч, а эукариоты обычно кодируют от нескольких тысяч до десятков тысяч белков (см. размер генома список примеров).
Биохимия

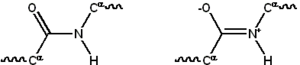
Большинство белков состоит из линейных полимеры построено из серий до 20 различных L-α- аминокислоты. Все протеиногенные аминокислоты обладают общими структурными особенностями, в том числе α-углерод к которому амино- группа, а карбоксил группа и переменная боковая цепь находятся связанный. Только пролин отличается от этой основной структуры, поскольку она содержит необычное кольцо для N-концевой аминогруппы, которое заставляет амидный фрагмент CO – NH принимать фиксированную конформацию.[23] Боковые цепи стандартных аминокислот, подробно описанные в список стандартных аминокислот, обладают большим разнообразием химических структур и свойств; именно комбинированное действие всех боковых цепей аминокислот в белке в конечном итоге определяет его трехмерную структуру и его химическую реакционную способность.[24]Аминокислоты в полипептидной цепи связаны между собой пептидные связи. После связывания в белковой цепи отдельная аминокислота называется остаток и связанный ряд атомов углерода, азота и кислорода известен как основная цепь или же белковый каркас.[25]
Пептидная связь имеет два резонанс формы, которые вносят некоторые двойная связь характер и запрещают вращение вокруг своей оси, так что альфа-атомы углерода примерно копланарный. Два других двугранные углы в пептидной связи определяют локальную форму, которую принимает остов белка.[26] Конец со свободной аминогруппой известен как N-конец или амино-конец, тогда как конец белка со свободной карбоксильной группой известен как C-конец или карбокси-конец (последовательность белка записывается от N-конца до C-конца слева направо).
Слова белок, полипептид, и пептид немного двусмысленны и могут совпадать по смыслу. Протеин обычно используется для обозначения полной биологической молекулы в стабильной конформация, в то время как пептид обычно используется для олигомеров с короткими аминокислотами, часто не имеющих стабильной трехмерной структуры. Но граница между ними четко не определена и обычно проходит около 20–30 остатков.[27] Полипептид может относиться к любой одиночной линейной цепи аминокислот, обычно независимо от длины, но часто подразумевает отсутствие определенного конформация.
Взаимодействия
Белки могут взаимодействовать со многими типами молекул, включая с другими белками, с липидами, с углеводами, и с ДНК.[28][29][30][31]
Изобилие в клетках
Было подсчитано, что средний размер бактерии содержат около 2 миллионов белков на клетку (например, Кишечная палочка и Золотистый стафилококк ). Более мелкие бактерии, такие как Микоплазма или же спирохеты содержат меньше молекул, от 50 000 до 1 миллиона. Напротив, эукариотический клетки больше и поэтому содержат намного больше белка. Например, дрожжи клетки, по оценкам, содержат около 50 миллионов белков и человек ячеек порядка от 1 до 3 миллиардов.[32] Концентрация отдельных белковых копий колеблется от нескольких молекул на клетку до 20 миллионов.[33] Не все гены, кодирующие белки, экспрессируются в большинстве клеток, и их количество зависит, например, от типа клетки и внешних стимулов. Например, из примерно 20 000 белков, кодируемых геномом человека, только 6000 обнаруживаются в лимфобластоидный клетки.[34]
Синтез
Биосинтез

Белки собираются из аминокислот с использованием информации, закодированной в генах. Каждый белок имеет свою уникальную аминокислотную последовательность, которая указана нуклеотид последовательность гена, кодирующего этот белок. В генетический код представляет собой набор из трех нуклеотидных наборов, называемых кодоны и каждая комбинация из трех нуклеотидов обозначает аминокислоту, например AUG (аденин –урацил –гуанин ) - это код для метионин. Поскольку ДНК содержит четыре нуклеотида, общее количество возможных кодонов составляет 64; следовательно, в генетическом коде присутствует некоторая избыточность, причем некоторые аминокислоты определяются более чем одним кодоном.[35] Гены, закодированные в ДНК, в первую очередь записано в пре-информационная РНК (мРНК) белками, такими как РНК-полимераза. Большинство организмов затем обрабатывают пре-мРНК (также известную как первичная стенограмма) с использованием различных форм Посттранскрипционная модификация для формирования зрелой мРНК, которая затем используется в качестве матрицы для синтеза белка рибосома. В прокариоты мРНК может быть использована сразу после ее образования или связана с рибосомой после того, как она отошла от нуклеоид. В отличие, эукариоты сделать мРНК в ядро клетки а потом перемещать это через ядерная мембрана в цитоплазма, куда синтез белка затем имеет место. Скорость синтеза белка у прокариот выше, чем у эукариот, и может достигать до 20 аминокислот в секунду.[36]
Процесс синтеза белка из матрицы мРНК известен как перевод. МРНК загружается на рибосому и считывается с трех нуклеотидов за раз путем сопоставления каждого кодона с его базовая пара антикодон расположен на переносить РНК молекула, которая несет аминокислоту, соответствующую распознаваемому кодону. Фермент аминоацил тРНК синтетаза «заряжает» молекулы тРНК правильными аминокислотами. Растущий полипептид часто называют зарождающаяся цепь. Белки всегда биосинтезируются из N-конец к C-конец.[35]
Размер синтезируемого белка можно измерить по количеству содержащихся в нем аминокислот и по его общему количеству. молекулярная масса, который обычно указывается в единицах дальтон (синоним атомные единицы массы ) или производной единицы килодальтон (кДа). Средний размер белка увеличивается от архей к бактериям и эукариотам (283, 311, 438 остатков и 31, 34, 49 кДа соответственно) из-за большего количества белковые домены составляющие белки у высших организмов.[37] Например, дрожжи белки состоят в среднем из 466 аминокислот в длину и 53 кДа по массе.[27] Самые большие известные белки - это тайтины, компонент мышца саркомер, с молекулярной массой почти 3000 кДа и общей длиной почти 27000 аминокислот.[38]
Химический синтез
Короткие белки также можно синтезировать химическим путем с помощью семейства методов, известных как пептидный синтез, которые полагаются на органический синтез методы, такие как химическая перевязка для получения пептидов с высоким выходом.[39] Химический синтез позволяет вводить неприродные аминокислоты в полипептидные цепи, такие как присоединение флуоресцентный зонды боковых цепей аминокислот.[40] Эти методы полезны в лаборатории. биохимия и клеточная биология, хотя обычно не для коммерческих приложений. Химический синтез неэффективен для полипептидов, длина которых превышает примерно 300 аминокислот, и синтезированные белки не могут легко принять свои природные третичная структура. Большинство методов химического синтеза идут от С-конца к N-концу, в противоположность биологической реакции.[41]
Структура

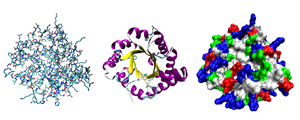
Большинство белков складывать в уникальные 3D-структуры. Форма, в которую естественным образом сворачивается белок, известна как его родное телосложение.[42] Хотя многие белки могут сворачиваться без посторонней помощи, просто благодаря химическим свойствам их аминокислот, другим требуется помощь молекулярных шапероны сворачиваться в свои родные состояния.[43] Биохимики часто называют четыре различных аспекта структуры белка:[44]
- Первичная структура: the аминокислотная последовательность. Белок - это полиамид.
- Вторичная структура: регулярно повторяющиеся локальные структуры, стабилизированные водородные связи. Наиболее распространенными примерами являются α-спираль, β-лист и повороты. Поскольку вторичные структуры являются локальными, в одной и той же молекуле белка может присутствовать множество областей разной вторичной структуры.
- Третичная структура: общая форма отдельной белковой молекулы; пространственное отношение вторичных структур друг к другу. Третичная структура обычно стабилизируется нелокальными взаимодействиями, чаще всего образованием гидрофобное ядро, но и через соляные мосты, водородные связи, дисульфидные связи, и даже посттрансляционные модификации. Термин «третичная структура» часто используется как синоним термина складывать. Третичная структура - это то, что контролирует основную функцию белка.
- Четвертичная структура: структура, образованная несколькими белковыми молекулами (полипептидными цепями), обычно называемая белковые субъединицы в этом контексте, которые функционируют как единый белковый комплекс.
- Пятеричная структура: признаки поверхности белка, которые организуют переполненный клеточный интерьер. Пятеричная структура зависит от временных, но существенных макромолекулярных взаимодействий, которые происходят внутри живых клеток.
Белки - это не совсем жесткие молекулы. В дополнение к этим уровням структуры белки могут перемещаться между несколькими родственными структурами, пока они выполняют свои функции. В контексте этих функциональных перестроек эти третичные или четвертичные структуры обычно называют "конформации ", а переходы между ними называются конформационные изменения. Такие изменения часто вызваны связыванием субстрат молекула фермента активный сайт, или физическая область белка, которая участвует в химическом катализе. В растворе белки также претерпевают изменения в структуре из-за тепловых колебаний и столкновений с другими молекулами.[45]

Белки можно неформально разделить на три основных класса, которые соотносятся с типичными третичными структурами: глобулярные белки, волокнистые белки, и мембранные белки. Почти все глобулярные белки растворимый и многие из них - ферменты. Волокнистые белки часто бывают структурными, например коллаген, основной компонент соединительной ткани, или кератин, белковый компонент волос и ногтей. Мембранные белки часто служат рецепторы или обеспечить каналы для полярных или заряженных молекул, чтобы пройти через клеточная мембрана.[46]
Частный случай внутримолекулярных водородных связей внутри белков, плохо защищенных от атаки воды и, следовательно, способствующих возникновению собственных обезвоживание, называются дегидроны.[47]
Белковые домены
Многие белки состоят из нескольких белковые домены, то есть сегменты белка, которые складываются в отдельные структурные единицы. Домены обычно также имеют определенные функции, такие как ферментативный деятельность (например, киназа ) или они служат в качестве связывающих модулей (например, SH3 домен связывается с богатыми пролином последовательностями других белков).
Мотив последовательности
Короткие аминокислотные последовательности в белках часто действуют как сайты узнавания для других белков.[48] Например, SH3 домены обычно связываются с короткими мотивами PxxP (т.е. 2 пролины [P], разделенные двумя неопределенными аминокислоты [x], хотя окружающие аминокислоты могут определять точную специфичность связывания). Многие из таких мотивов собраны в Эукариотический линейный мотив (ELM) база данных.
Клеточные функции
Белки - главные действующие лица клетки, которые, как говорят, выполняют функции, определенные информацией, закодированной в генах.[27] За исключением некоторых типов РНК, большинство других биологических молекул являются относительно инертными элементами, на которые действуют белки. Белки составляют половину сухой массы кишечная палочка клетка, тогда как другие макромолекулы, такие как ДНК и РНК, составляют только 3% и 20% соответственно.[49] Набор белков, экспрессируемых в определенных клетках или типах клеток, известен как их протеом.

Основная характеристика белков, которая также обеспечивает их разнообразный набор функций, - это их способность специфически и прочно связывать другие молекулы. Область белка, отвечающая за связывание другой молекулы, известна как сайт привязки и часто представляет собой углубление или «карман» на молекулярной поверхности. Эта связывающая способность опосредована третичной структурой белка, которая определяет карман сайта связывания, и химическими свойствами боковых цепей окружающих аминокислот. Связывание с белками может быть чрезвычайно прочным и специфичным; например, ингибитор рибонуклеазы белок связывается с человеком ангиогенин с субфемтомолярным константа диссоциации (<10−15 M), но совершенно не связывается со своим гомологом амфибии онконасе (> 1 млн). Чрезвычайно незначительные химические изменения, такие как добавление одной метильной группы к связывающему партнеру, иногда могут быть достаточными для почти полного устранения связывания; например, аминоацил тРНК синтетаза специфичен для аминокислоты валин различает очень похожую боковую цепь аминокислоты изолейцин.[50]
Белки могут связываться с другими белками, а также с малая молекула субстраты. Когда белки специфически связываются с другими копиями той же молекулы, они могут олигомеризовать образовывать фибриллы; этот процесс часто происходит в структурных белках, которые состоят из глобулярных мономеров, которые самоассоциируются с образованием жестких волокон. Белковые взаимодействия также регулируют ферментативную активность, контролируют прогрессирование через клеточный цикл, и позволяют сборку больших белковые комплексы которые выполняют множество тесно связанных реакций с общей биологической функцией. Белки также могут связываться с клеточными мембранами или даже интегрироваться в них. Способность партнеров по связыванию вызывать конформационные изменения в белках позволяет создавать чрезвычайно сложные сигнализация сети.[51]Поскольку взаимодействия между белками обратимы и в значительной степени зависят от доступности различных групп белков-партнеров для образования агрегатов, способных выполнять дискретные наборы функций, изучение взаимодействий между конкретными белками является ключом к пониманию важных аспектов клеточной функции. и, в конечном счете, свойства, которые различают определенные типы клеток.[52][53]
Ферменты
Наиболее известная роль белков в клетке заключается в следующем. ферменты, который катализировать химические реакции. Ферменты обычно очень специфичны и ускоряют только одну или несколько химических реакций. Ферменты проводят большинство реакций, связанных с метаболизм, а также манипулирование ДНК в таких процессах, как Репликация ДНК, Ремонт ДНК, и транскрипция. Некоторые ферменты воздействуют на другие белки, добавляя или удаляя химические группы в процессе, известном как посттрансляционная модификация. Известно, что ферменты катализируют около 4000 реакций.[54] Ускорение скорости, обеспечиваемое ферментативным катализом, часто бывает огромным - до 1017-кратное увеличение скорости по сравнению с некаталитической реакцией в случае оротат декарбоксилаза (78 миллионов лет без фермента, 18 миллисекунд с ферментом).[55]
Молекулы, связанные ферментами и действующие на них, называются субстраты. Хотя ферменты могут состоять из сотен аминокислот, обычно только небольшая часть остатков контактирует с субстратом, а еще меньшая часть - в среднем от трех до четырех остатков - непосредственно участвует в катализе.[56] Область фермента, которая связывает субстрат и содержит каталитические остатки, известна как активный сайт.
Диригентные белки являются членами класса белков, которые диктуют стереохимия соединения, синтезируемого другими ферментами.[57]
Передача клеточных сигналов и связывание лиганда
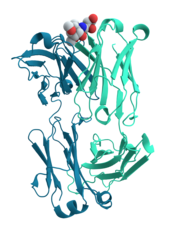
Многие белки участвуют в процессе клеточная сигнализация и преобразование сигнала. Некоторые белки, такие как инсулин, являются внеклеточными белками, которые передают сигнал от клетки, в которой они были синтезированы, другим клеткам в отдаленных ткани. Другие мембранные белки которые действуют как рецепторы чья основная функция - связывать сигнальную молекулу и вызывать биохимический ответ в клетке. Многие рецепторы имеют сайт связывания, расположенный на поверхности клетки, и эффекторный домен внутри клетки, который может обладать ферментативной активностью или может подвергаться действию конформационное изменение обнаруживается другими белками внутри клетки.[58]
Антитела белковые компоненты адаптивная иммунная система чья основная функция - связывать антигены, или посторонние вещества в организме, и нацелить их на разрушение. Антитела могут быть секретный во внеклеточную среду или закреплены в мембранах специализированных В-клетки известный как плазматические клетки. В то время как ферменты ограничены в их аффинности связывания со своими субстратами из-за необходимости проведения реакции, антитела не имеют таких ограничений. Аффинность связывания антитела с его мишенью чрезвычайно высока.[59]
Многие транспортные белки лигандов связывают определенные маленькие биомолекулы и транспортировать их в другие места в организме многоклеточного организма. Эти белки должны иметь высокую аффинность связывания, когда их лиганд присутствует в высоких концентрациях, но также должен высвобождать лиганд, когда он присутствует в низких концентрациях в тканях-мишенях. Каноническим примером лиганд-связывающего белка является гемоглобин, который транспортирует кислород от легкие к другим органам и тканям во всех позвоночные и имеет близких гомологов во всех биологических Королевство.[60] Лектины находятся сахар-связывающие белки которые очень специфичны для своих сахарных фрагментов. Лектины обычно играют роль в биологических признание явления с участием клеток и белков.[61] Рецепторы и гормоны представляют собой высокоспецифичные связывающие белки.
Трансмембранные белки могут также служить белками-транспортерами лигандов, которые изменяют проницаемость клеточной мембраны маленькие молекулы и ионы. Одна только мембрана имеет гидрофобный ядро, через которое полярный или заряженные молекулы не могут размытый. Мембранные белки содержат внутренние каналы, которые позволяют таким молекулам входить и выходить из клетки. Много ионный канал белки специализируются на выборе только определенного иона; Например, калий и натрий каналы часто различают только один из двух ионов.[62]
Структурные белки
Структурные белки придают жесткость и жесткость биологическим компонентам, которые иначе текли. Большинство структурных белков волокнистые белки; Например, коллаген и эластин являются важнейшими компонентами соединительная ткань Такие как хрящ, и кератин встречается в твердых или нитчатых структурах, таких как волосы, гвозди, перья, копыта, и немного панцири животных.[63] Немного глобулярные белки может также выполнять структурные функции, например, актин и тубулин глобулярны и растворимы как мономеры, но полимеризовать образовывать длинные жесткие волокна, из которых состоит цитоскелет, что позволяет клетке сохранять свою форму и размер.
Другие белки, которые выполняют структурные функции: моторные белки Такие как миозин, кинезин, и динеин, которые способны создавать механические силы. Эти белки имеют решающее значение для клеточного подвижность одноклеточных организмов и сперма многих многоклеточных организмов, которые воспроизводят сексуально. Они также создают силы, возникающие при сокращении мышцы[64] и играют важную роль во внутриклеточном транспорте.
Методы исследования
Можно исследовать активность и структуру белков. in vitro, in vivo, и in silico. В пробирке исследования очищенных белков в контролируемой среде полезны для изучения того, как белок выполняет свою функцию: например, кинетика ферментов исследования исследуют химический механизм каталитической активности фермента и его относительного сродства к различным возможным молекулам субстрата. Напротив, in vivo эксперименты могут предоставить информацию о физиологической роли белка в контексте клетка или даже целый организм. In silico исследования используют вычислительные методы для изучения белков.
Очистка белков
Выполнить in vitro анализ, белок должен быть очищен от других клеточных компонентов. Этот процесс обычно начинается с лизис клеток, при котором мембрана клетки разрушается, и ее внутреннее содержимое выделяется в раствор, известный как сырой лизат. Полученную смесь можно очистить, используя ультрацентрифугирование, который фракционирует различные клеточные компоненты на фракции, содержащие растворимые белки; мембрана липиды и белки; сотовый органеллы, и нуклеиновые кислоты. Осадки методом, известным как высаливание может концентрировать белки из этого лизата. Различные виды хроматография затем используются для выделения интересующего белка или белков на основе таких свойств, как молекулярная масса, чистый заряд и сродство связывания.[65] Уровень очистки можно контролировать с помощью различных типов гель-электрофорез если желаемый молекулярный вес белка и изоэлектрическая точка известны по спектроскопия если белок имеет различимые спектроскопические особенности, или ферментные анализы если белок обладает ферментативной активностью. Кроме того, белки можно выделить в соответствии с их зарядом, используя электрофокусировка.[66]
Для натуральных белков может потребоваться серия этапов очистки для получения белка, достаточно чистого для лабораторных применений. Чтобы упростить этот процесс, генная инженерия часто используется для добавления к белкам химических свойств, которые упрощают их очистку, не влияя на их структуру или активность. Здесь «метка», состоящая из определенной аминокислотной последовательности, часто серии гистидин остатки (а "Его тег "), присоединяется к одному концу белка. В результате, когда лизат пропускается через хроматографическую колонку, содержащую никель остатки гистидина связывают никель и прикрепляются к колонке, в то время как немаркированные компоненты лизата проходят беспрепятственно. Был разработан ряд различных тегов, чтобы помочь исследователям очистить определенные белки от сложных смесей.[67]
Сотовая локализация
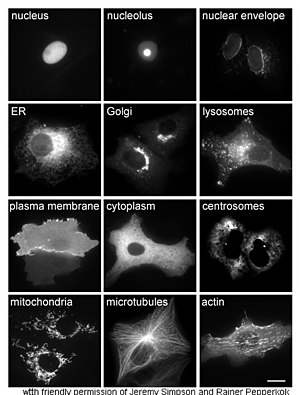
Изучение белков in vivo часто занимается синтезом и локализацией белка в клетке. Хотя многие внутриклеточные белки синтезируются в цитоплазма и мембраносвязанные или секретируемые белки в эндоплазматический ретикулум, особенности того, как белки целевой к конкретным органеллам или клеточным структурам часто неясно. Полезный метод оценки клеточной локализации использует генную инженерию для экспрессии в клетке гибридный белок или же химера состоящий из интересующего природного белка, связанного с "репортер " Такие как зеленый флуоресцентный белок (GFP).[68] Положение слитого белка в клетке можно четко и эффективно визуализировать с помощью микроскопия,[69] как показано на рисунке напротив.
Другие методы выяснения клеточного местоположения белков требуют использования известных компартментных маркеров для таких областей, как ER, Гольджи, лизосомы или вакуоли, митохондрии, хлоропласты, плазматическая мембрана и т. Д. С использованием флуоресцентно меченных версий этих маркеров или антител к известным маркерам становится намного проще определить локализацию интересующего белка. Например, непрямая иммунофлуоресценция позволит колокализацию флуоресценции и демонстрацию местоположения. Флуоресцентные красители используются для маркировки клеточных компартментов с аналогичной целью.[70]
Существуют и другие возможности. Например, иммуногистохимия обычно используют антитело к одному или нескольким интересующим белкам, которые конъюгированы с ферментами, дающими люминесцентные или хромогенные сигналы, которые можно сравнивать между образцами, что позволяет получить информацию о локализации. Другой применимый метод - это совместное фракционирование в градиентах сахарозы (или другого материала) с использованием изопикническое центрифугирование.[71] Хотя этот метод не доказывает совместную локализацию компартмента известной плотности и интересующего белка, он увеличивает вероятность и более поддается крупномасштабным исследованиям.
Наконец, золотой стандарт клеточной локализации - это иммуноэлектронная микроскопия. В этом методе также используются антитела к интересующему белку, наряду с классическими методами электронной микроскопии. Образец готовят для обычного исследования под электронным микроскопом, а затем обрабатывают антителом к интересующему белку, который конъюгирован с чрезвычайно электроплотным материалом, обычно золотом. Это позволяет локализовать как детали ультраструктуры, так и интересующий белок.[72]
Через другое приложение генной инженерии, известное как сайт-направленный мутагенез, исследователи могут изменять последовательность белка и, следовательно, его структуру, клеточную локализацию и чувствительность к регуляции. Этот метод позволяет даже включать неприродные аминокислоты в белки с использованием модифицированных тРНК,[73] и может позволить рациональное дизайн новых белков с новыми свойствами.[74]
Протеомика
Общий набор белков, одновременно присутствующих в клетке или типе клеток, известен как их протеом, и изучение таких крупномасштабных наборов данных определяет область протеомика, названный по аналогии с родственным полем геномика. Ключевые экспериментальные методы в протеомике включают: 2D электрофорез,[75] который позволяет разделить множество белков, масс-спектрометрии,[76] что позволяет быстро идентифицировать белки и секвенировать пептиды с высокой пропускной способностью (чаще всего после переваривание в геле ), белковые микрочипы, которые позволяют определять относительные уровни различных белков, присутствующих в клетке, и двухгибридный скрининг, что позволяет систематически исследовать белок-белковые взаимодействия.[77] Полный набор биологически возможных таких взаимодействий известен как интерактом.[78] Систематическая попытка определить структуры белков, представляющих все возможные складки, известна как структурная геномика.[79]
Биоинформатика
Для анализа структуры, функции и эволюции белков был разработан широкий спектр вычислительных методов.
Разработка таких инструментов была вызвана большим количеством геномных и протеомных данных, доступных для различных организмов, включая человеческий геном. Экспериментально изучить все белки просто невозможно, поэтому только некоторые из них подвергаются лабораторным экспериментам, в то время как вычислительные инструменты используются для экстраполяции на похожие белки. Такой гомологичные белки могут быть эффективно идентифицированы у отдаленно родственных организмов с помощью выравнивание последовательностей. Геном и последовательности генов можно искать с помощью различных инструментов на предмет определенных свойств. Инструменты профилирования последовательности могу найти рестрикционный фермент места, открытые рамки для чтения в нуклеотид последовательности и предсказывать второстепенные конструкции. Филогенетические деревья могут быть построены и эволюционный гипотезы, разработанные с помощью специального программного обеспечения, такого как ClustalW относительно происхождения современных организмов и генов, которые они выражают. Поле биоинформатика теперь незаменим для анализа генов и белков.
Определение структуры
Обнаружение третичной структуры белка или четвертичной структуры его комплексов может дать важные подсказки о том, как белок выполняет свою функцию и как на него можно повлиять, т.е. дизайн препарата. Как белки слишком маленький, чтобы его можно было увидеть под оптический микроскоп необходимо использовать другие методы для определения их структуры. Общие экспериментальные методы включают: Рентгеновская кристаллография и ЯМР-спектроскопия, оба из которых могут производить структурную информацию на атомный разрешающая способность. Однако эксперименты с ЯМР могут предоставить информацию, на основе которой можно оценить подмножество расстояний между парами атомов, а окончательные возможные конформации для белка определяются путем решения геометрия расстояния проблема. Двойная поляризационная интерферометрия это количественный аналитический метод измерения общей конформация белка и конформационные изменения из-за взаимодействий или других стимулов. Круговой дихроизм это еще один лабораторный метод определения внутреннего β-листового / α-спирального состава белков. Криоэлектронная микроскопия используется для получения структурной информации с низким разрешением об очень больших белковых комплексах, в том числе собранных вирусы;[80] вариант, известный как электронная кристаллография в некоторых случаях может также давать информацию с высоким разрешением, особенно для двумерных кристаллов мембранных белков.[81] Решенные структуры обычно откладываются в Банк данных белков (PDB), свободно доступный ресурс, из которого можно получить структурные данные о тысячах белков в виде Декартовы координаты для каждого атома в белке.[82]
Известно гораздо больше генных последовательностей, чем белковых структур. Кроме того, набор решенных структур смещен в сторону белков, которые можно легко подвергнуть условиям, необходимым в Рентгеновская кристаллография, один из основных методов определения структуры. В частности, глобулярные белки сравнительно легко кристаллизоваться при подготовке к рентгеновской кристаллографии. Мембранные белки и крупные белковые комплексы, напротив, трудно кристаллизовать, и они недостаточно представлены в PDB.[83] Структурная геномика инициативы пытались исправить эти недостатки путем систематического решения репрезентативных структур основных классов складок. Прогноз структуры белка Методы пытаются предоставить средства создания вероятной структуры для белков, структуры которых не были определены экспериментально.[84]
Прогнозирование и моделирование конструкции

Дополняет область структурной геномики, предсказание структуры белка развивается эффективный математические модели белков для теоретического предсказания молекулярных образований с помощью вычислений вместо обнаружения структур с помощью лабораторных наблюдений.[85] Самый успешный тип предсказания структуры, известный как моделирование гомологии, основывается на существовании «шаблонной» структуры с последовательностью, подобной моделируемому белку; Цель структурной геномики - обеспечить достаточное представление в решенных структурах для моделирования большинства из оставшихся.[86] Хотя создание точных моделей остается проблемой, когда доступны только отдаленно связанные структуры шаблонов, было высказано предположение, что выравнивание последовательностей является узким местом в этом процессе, поскольку достаточно точные модели могут быть получены, если известно «идеальное» выравнивание последовательностей.[87] Многие методы прогнозирования структуры служат источником информации для развивающейся области белковая инженерия, в котором уже созданы новые белковые складки.[88] Более сложная вычислительная проблема - это предсказание межмолекулярных взаимодействий, например, в молекулярный док и прогноз межбелкового взаимодействия.[89]
Математические модели для моделирования динамических процессов сворачивание белка и связывание включает молекулярная механика, особенно, молекулярная динамика. Монте-Карло методы облегчают вычисления, которые используют достижения параллельно и распределенных вычислений (например, Складной @ дома проект[90] который выполняет молекулярное моделирование на GPU ). In silico моделирование обнаружило сворачивание небольших α-спиральных белковые домены такой как Виллин головной убор[91] и ВИЧ вспомогательный белок.[92] Гибридные методы, сочетающие стандартную молекулярную динамику с квантово-механический математика исследовала электронные состояния родопсины.[93]
Прогнозирование белковых нарушений и неструктурирования
Многие белки (у эукариот ~ 33%) содержат большие неструктурированные, но биологически функциональные сегменты и могут быть классифицированы как внутренне неупорядоченные белки.[94] Таким образом, прогнозирование и анализ нарушения белков является важной частью характеристики структуры белка.[95]
Питание
Наиболее микроорганизмы и растения могут биосинтезировать все 20 стандартных аминокислоты, в то время как животные (включая человека) должны получать некоторые аминокислоты из рацион питания.[49] Аминокислоты, которые организм не может синтезировать самостоятельно, называются незаменимые аминокислоты. Ключевые ферменты, которые синтезируют определенные аминокислоты, отсутствуют у животных, например аспартокиназа, который катализирует первую стадию синтеза лизин, метионин, и треонин из аспартат. Если в окружающей среде присутствуют аминокислоты, микроорганизмы могут сохранять энергию, поглощая аминокислоты из окружающей среды и подавление их биосинтетические пути.
У животных аминокислоты получают в результате потребления продуктов, содержащих белок. Затем проглоченные белки расщепляются на аминокислоты через пищеварение, который обычно включает денатурация белка в результате воздействия кислота и гидролиз ферментами, называемыми протеазы. Некоторые поступившие аминокислоты используются для биосинтеза белка, а другие превращаются в глюкоза через глюконеогенез, или подается в цикл лимонной кислоты. Использование белка в качестве топлива особенно важно при голодание условиях, поскольку он позволяет использовать собственные белки организма для поддержания жизни, особенно те, которые содержатся в мышца.[96]
У таких животных, как собаки и кошки, белок поддерживает здоровье и качество кожи, способствуя росту волосяных фолликулов и кератинизации, тем самым снижая вероятность проблем с кожей, вызывающих неприятный запах.[97] Низкокачественные белки также влияют на здоровье желудочно-кишечного тракта, увеличивая вероятность метеоризма и пахучих соединений у собак, потому что, когда белки достигают толстой кишки в непереваренном состоянии, они ферментируются с образованием сероводорода, индола и скатола.[98] Собаки и кошки переваривают животные белки лучше, чем растительные, но продукты низкого качества животного происхождения, в том числе кожа, перья и соединительная ткань, перевариваются плохо.[98]
Химический анализ
Общий азот органического вещества в основном формируется аминогруппами белков. Общий азот по Кьельдалю (ТКН ) является показателем азота, широко используемым при анализе (сточных) воды, почвы, пищевых продуктов, кормов и органических веществ в целом. Как следует из названия, Метод Кьельдаля применяется. Доступны более чувствительные методы [99][100].
Смотрите также
Рекомендации
- ^ Томас Бёрр Осборн (1909): Растительные белки В архиве 2016-03-22 в Wayback Machine, История с 1 по 6, с archive.org
- ^ Малдер GJ (1838). "Sur la композиция из живых веществ". Bulletin des Sciences Physiques et Naturelles en Néerlande: 104.
- ^ Гарольд Х (1951). «Происхождение слова« белок ».'". Природа. 168 (4267): 244. Bibcode:1951Натура.168..244H. Дои:10.1038 / 168244a0. PMID 14875059. S2CID 4271525.
- ^ а б c Перретт Д. (август 2007 г.). «От« протеина »к истокам клинической протеомики». Протеомика: клиническое применение. 1 (8): 720–38. Дои:10.1002 / prca.200700525. PMID 21136729. S2CID 32843102.
- ^ Новый оксфордский словарь английского языка
- ^ Рейнольдс Дж. А., Танфорд С. (2003). Роботы природы: история белков (Оксфордские книги в мягкой обложке). Нью-Йорк, Нью-Йорк: Издательство Оксфордского университета. п. 15. ISBN 978-0-19-860694-9.
- ^ Рейнольдс и Танфорд (2003).
- ^ Бишофф Т.Л., Войт С (1860). Die Gesetze der Ernaehrung des Pflanzenfressers durch neue Untersuchungen festgestellt (на немецком). Лейпциг, Гейдельберг.
- ^ "Хофмайстер, Франц". encyclopedia.com. В архиве из оригинала 5 апреля 2017 г.. Получено 4 апреля 2017.
- ^ «Белок, раздел: Классификация белков». britannica.com. В архиве из оригинала 4 апреля 2017 г.. Получено 4 апреля 2017.
- ^ Самнер Дж. Б. (1926). «Выделение и кристаллизация фермента уреазы. Предварительная статья» (PDF). Журнал биологической химии. 69 (2): 435–41. В архиве из оригинала 2011-03-25. Получено 2011-01-16.
- ^ Полинг Л., Кори Р. Б. (май 1951 г.). «Координаты атомов и структурные факторы для двух спиральных конфигураций полипептидных цепей» (PDF). Труды Национальной академии наук Соединенных Штатов Америки. 37 (5): 235–40. Bibcode:1951ПНАС ... 37..235П. Дои:10.1073 / pnas.37.5.235. ЧВК 1063348. PMID 14834145. В архиве (PDF) из оригинала от 28.11.2012. Получено 2009-04-14.
- ^ Каузманн В. (май 1956 г.). «Структурные факторы денатурации белков». Журнал клеточной физиологии. 47 (Дополнение 1): 113–31. Дои:10.1002 / jcp.1030470410. PMID 13332017.
- ^ Каузманн В. (1959). «Некоторые факторы в интерпретации денатурации белков». Достижения в химии белков Том 14. Успехи в химии белков. 14. С. 1–63. Дои:10.1016 / S0065-3233 (08) 60608-7. ISBN 978-0-12-034214-3. PMID 14404936.
- ^ Кальман С.М., Линдерстрём-Ланг К., Оттесен М., Ричардс Ф.М. (февраль 1955 г.). «Расщепление рибонуклеазы субтилизином». Biochimica et Biophysica Acta. 16 (2): 297–99. Дои:10.1016/0006-3002(55)90224-9. PMID 14363272.
- ^ Сэнгер Ф (1949). «Концевые пептиды инсулина». Биохимический журнал. 45 (5): 563–74. Дои:10.1042 / bj0450563. ЧВК 1275055. PMID 15396627.
- ^ Сэнгер Ф. (1958), Нобелевская лекция: химия инсулина (PDF), Nobelprize.org, в архиве (PDF) из оригинала от 05.01.2013, получено 2016-02-09
- ^ Muirhead H, Perutz MF (август 1963 г.). «Структура гемоглобина. Трехмерный синтез Фурье восстановленного гемоглобина человека с разрешением 5,5 Å». Природа. 199 (4894): 633–38. Bibcode:1963Натура.199..633М. Дои:10.1038 / 199633a0. PMID 14074546. S2CID 4257461.
- ^ Кендрю Дж. К., Бодо Дж., Динцис Х. М., Пэрриш Р. Г., Вайкофф Х., Филлипс, округ Колумбия (март 1958 г.). «Трехмерная модель молекулы миоглобина, полученная методом рентгеновского анализа». Природа. 181 (4610): 662–66. Bibcode:1958Натура.181..662K. Дои:10.1038 / 181662a0. PMID 13517261. S2CID 4162786.
- ^ «RCSB Protein Data Bank». Архивировано из оригинал на 2015-04-18. Получено 2017-01-19.
- ^ Чжоу Чж (апрель 2008 г.). «К атомному разрешению структурного определения с помощью одночастичной криоэлектронной микроскопии». Текущее мнение в структурной биологии. 18 (2): 218–28. Дои:10.1016 / j.sbi.2008.03.004. ЧВК 2714865. PMID 18403197.
- ^ Кескин О., Танцбаг Н., Гурсой А. (апрель 2008 г.). «Характеристика и прогнозирование белковых интерфейсов для вывода сетей белок-белкового взаимодействия». Текущая фармацевтическая биотехнология. 9 (2): 67–76. Дои:10.2174/138920108783955191. PMID 18393863.
- ^ Нельсон Д.Л., Кокс М.М. (2005). Принципы биохимии Ленингера (4-е изд.). Нью-Йорк, Нью-Йорк: В. Х. Фриман и компания.
- ^ Гаттеридж А., Торнтон Дж. М. (ноябрь 2005 г.). «Понимание каталитического инструментария природы». Тенденции в биохимических науках. 30 (11): 622–29. Дои:10.1016 / j.tibs.2005.09.006. PMID 16214343.
- ^ Мюррей и другие., п. 19.
- ^ Мюррей и другие., п. 31.
- ^ а б c Лодиш Х, Берк А., Мацудаира П., Кайзер Калифорния, Кригер М., Скотт М. П., Зипуркси С. Л., Дарнелл Дж. (2004). Молекулярная клеточная биология (5-е изд.). Нью-Йорк, Нью-Йорк: WH Freeman and Company.
- ^ Ardejani, Maziar S .; Пауэрс, Эван Т .; Келли, Джеффри В. (2017). «Использование кооперативно свернутых пептидов для измерения энергии взаимодействия и конформационных склонностей». Отчеты о химических исследованиях. 50 (8): 1875–82. Дои:10.1021 / acs.accounts.7b00195. ISSN 0001-4842. ЧВК 5584629. PMID 28723063.
- ^ Бранден С., Туз Дж. (1999). Введение в структуру белка. Нью-Йорк: Garland Pub. ISBN 978-0-8153-2305-1.
- ^ Мюррей РФ, Харпер Х.В., Граннер Д.К., Мэйс ПА, Родвелл Ф.В. (2006). Иллюстрированная биохимия Харпера. Нью-Йорк: Lange Medical Books / McGraw-Hill. ISBN 978-0-07-146197-9.
- ^ Ван Холд К. Э., Мэтьюз К. К. (1996). Биохимия. Менло-Парк, Калифорния: Паб Бенджамин / Каммингс. Co., Inc. ISBN 978-0-8053-3931-4.
- ^ Майло Р. (декабрь 2013 г.). «Каково общее количество белковых молекул на объем клетки? Призыв переосмыслить некоторые опубликованные значения». BioEssays. 35 (12): 1050–55. Дои:10.1002 / bies.201300066. ЧВК 3910158. PMID 24114984.
- ^ Бек М., Шмидт А., Мальмстроем Дж., Клаассен М., Ори А., Шимборска А., Херцог Ф., Риннер О., Элленберг Дж., Эберсолд Р. (ноябрь 2011 г.). «Количественный протеом линии клеток человека». Молекулярная системная биология. 7: 549. Дои:10.1038 / msb.2011.82. ЧВК 3261713. PMID 22068332.
- ^ Ву Л., Кандилл С.И., Чой Й, Се Д., Цзян Л., Ли-Пок-Тхан Дж., Тан Х., Снайдер М. (июль 2013 г.). «Вариации и генетический контроль обилия белка у людей». Природа. 499 (7456): 79–82. Bibcode:2013Натура 499 ... 79Вт. Дои:10.1038 / природа12223. ЧВК 3789121. PMID 23676674.
- ^ а б ван Холд и Мэтьюз, стр. 1002–42.
- ^ Добсон CM (2000). «Природа и значение сворачивания белков». In Pain RH (ред.). Механизмы сворачивания белков. Оксфорд, Оксфордшир: Издательство Оксфордского университета. С. 1–28. ISBN 978-0-19-963789-8.
- ^ Козловский Л.П. (январь 2017 г.). «Proteome-pI: база данных изоэлектрических точек протеома». Исследования нуклеиновых кислот. 45 (D1): D1112 – D1116. Дои:10.1093 / нар / gkw978. ЧВК 5210655. PMID 27789699.
- ^ Фултон А.Б., Айзекс В.Б. (апрель 1991 г.). «Титин, огромный эластичный саркомерный белок, который, вероятно, играет важную роль в морфогенезе». BioEssays. 13 (4): 157–61. Дои:10.1002 / bies.950130403. PMID 1859393. S2CID 20237314.
- ^ Брукдорфер Т., Мардер О., Альберисио Ф. (февраль 2004 г.). «От производства пептидов в миллиграммах для исследований до многотонных количеств для лекарств будущего». Текущая фармацевтическая биотехнология. 5 (1): 29–43. Дои:10.2174/1389201043489620. PMID 14965208.
- ^ Шварцер Д., Коул П.А. (декабрь 2005 г.). «Полусинтез белков и выраженное лигирование белков: погоня за хвостом белка». Современное мнение в области химической биологии. 9 (6): 561–69. Дои:10.1016 / j.cbpa.2005.09.018. PMID 16226484.
- ^ Кент С.Б. (февраль 2009 г.). «Полный химический синтез белков». Обзоры химического общества. 38 (2): 338–51. Дои:10.1039 / b700141j. PMID 19169452.
- ^ Мюррей и другие., п. 36.
- ^ Мюррей и другие., п. 37.
- ^ Мюррей и другие., стр. 30–34.
- ^ ван Холд и Мэтьюз, стр. 368–75.
- ^ ван Холд и Мэтьюз, стр. 165–85.
- ^ Фернандес А., Скотт Р. (сентябрь 2003 г.). «Дегидрон: структурно закодированный сигнал взаимодействия белков». Биофизический журнал. 85 (3): 1914–28. Bibcode:2003BpJ .... 85.1914F. Дои:10.1016 / S0006-3495 (03) 74619-0. ЧВК 1303363. PMID 12944304.
- ^ Дэйви Н.Э., Ван Рой К., Уэзеритт Р.Дж., Тоедт Дж., Уяр Б., Альтенберг Б., Бадд А., Дилла Ф., Динкель Х., Гибсон Т.Дж. (январь 2012 г.). «Атрибуты коротких линейных мотивов». Молекулярные биосистемы. 8 (1): 268–81. Дои:10.1039 / c1mb05231d. PMID 21909575.
- ^ а б Voet D, Voet JG. (2004). Биохимия Том 1 3-е изд. Wiley: Хобокен, штат Нью-Джерси.
- ^ Шанкаранараянан Р., Морас Д. (2001). «Верность перевода генетического кода». Acta Biochimica Polonica. 48 (2): 323–35. Дои:10.18388 / abp.2001_3918. PMID 11732604.
- ^ ван Холд и Мэтьюз, стр. 830–49.
- ^ Копленд Дж. А., Шеффилд-Мур М., Колдзич-Живанович Н., Джентри С., Лампру Дж., Цорцату-Статопулу Ф., Зумпурлис В., Урбан Р. Дж., Влахопулос С. А. (июнь 2009 г.). «Рецепторы половых стероидов в дифференцировке скелета и эпителиальной неоплазии: возможно ли тканеспецифическое вмешательство?». BioEssays. 31 (6): 629–41. Дои:10.1002 / bies.200800138. PMID 19382224. S2CID 205469320.
- ^ Самарин С., Нусрат А. (январь 2009 г.). «Регулирование эпителиального апикального соединительного комплекса GTPases семейства Rho». Границы биологических наук. 14 (14): 1129–42. Дои:10.2741/3298. PMID 19273120.
- ^ Байроч А. (январь 2000 г.). «База данных ENZYME в 2000 году» (PDF). Исследования нуклеиновых кислот. 28 (1): 304–05. Дои:10.1093 / nar / 28.1.304. ЧВК 102465. PMID 10592255. Архивировано из оригинал (PDF) 1 июня 2011 г.
- ^ Радзичка А., Вольфенден Р. (январь 1995 г.). «Опытный фермент». Наука. 267 (5194): 90–3. Bibcode:1995Научный ... 267 ... 90R. Дои:10.1126 / science.7809611. PMID 7809611.
- ^ EBI External Services (2010-01-20). "Атлас каталитического сайта в Европейском институте биоинформатики". Ebi.ac.uk. В архиве из оригинала 20.06.2013. Получено 2011-01-16.
- ^ Пикель Б., Шаллер А. (октябрь 2013 г.). «Диригентные белки: молекулярные характеристики и потенциальные биотехнологические применения». Прикладная микробиология и биотехнология. 97 (19): 8427–38. Дои:10.1007 / s00253-013-5167-4. PMID 23989917. S2CID 1896003.
- ^ Бранден и Туз, стр. 251–81.
- ^ ван Холд и Мэтьюз, стр. 247–50.
- ^ ван Холд и Мэтьюз, стр.220–29.
- ^ Рюдигер Х., Зиберт Х.С., Солис Д., Хименес-Барберо Дж., Ромеро А., фон дер Лиет К. В., Диас-Мариньо Т., Габиус Х. Дж. (Апрель 2000 г.). «Медицинская химия, основанная на сахарном коде: основы лектинологии и экспериментальные стратегии с лектинами в качестве мишеней». Современная лекарственная химия. 7 (4): 389–416. Дои:10.2174/0929867003375164. PMID 10702616.
- ^ Бранден и Туз, стр. 232–34.
- ^ ван Холд и Мэтьюз, стр. 178–81.
- ^ ван Холд и Мэтьюз, стр. 258–64, 272.
- ^ Мюррей и другие., стр. 21–24.
- ^ Эй Дж, Пош А., Коэн А., Лю Н., Харберс А. (2008). «Фракционирование сложных белковых смесей методом жидкофазного изоэлектрического фокусирования». 2D-СТРАНИЦА: Подготовка проб и фракционирование. Методы молекулярной биологии. Методы молекулярной биологии ™. 424. стр.225–39. Дои:10.1007/978-1-60327-064-9_19. ISBN 978-1-58829-722-8. PMID 18369866.
- ^ Терпе К (январь 2003 г.). «Обзор слияния белков-меток: от молекулярных и биохимических основ до коммерческих систем». Прикладная микробиология и биотехнология. 60 (5): 523–33. Дои:10.1007 / s00253-002-1158-6. PMID 12536251. S2CID 206934268.
- ^ Степаненко О.В., Верхуша В.В., Кузнецова И.М., Уверский В.Н., Туроверов К.К. (август 2008 г.). «Флуоресцентные белки как биомаркеры и биосенсоры: бросая цветные огни на молекулярные и клеточные процессы». Современная наука о белках и пептидах. 9 (4): 338–69. Дои:10.2174/138920308785132668. ЧВК 2904242. PMID 18691124.
- ^ Юсте Р. (декабрь 2005 г.). «Флуоресцентная микроскопия сегодня». Методы природы. 2 (12): 902–4. Дои:10.1038 / nmeth1205-902. PMID 16299474. S2CID 205418407.
- ^ Марголин В. (январь 2000 г.). «Зеленый флуоресцентный белок как репортер макромолекулярной локализации в бактериальных клетках». Методы. 20 (1): 62–72. Дои:10.1006 / мет.1999.0906. PMID 10610805.
- ^ Уокер Дж. Х., Уилсон К. (2000). Принципы и методы практической биохимии. Кембридж, Великобритания: Издательство Кембриджского университета. С. 287–89. ISBN 978-0-521-65873-7.
- ^ Mayhew TM, Lucocq JM (август 2008 г.). «Разработки в клеточной биологии для количественной иммуноэлектронной микроскопии на основе тонких срезов: обзор». Гистохимия и клеточная биология. 130 (2): 299–313. Дои:10.1007 / s00418-008-0451-6. ЧВК 2491712. PMID 18553098.
- ^ Хосака Т., Сисидо М. (декабрь 2002 г.). «Включение неприродных аминокислот в белки». Современное мнение в области химической биологии. 6 (6): 809–15. Дои:10.1016 / S1367-5931 (02) 00376-9. PMID 12470735.
- ^ Cedrone F, Ménez A, Quéméneur E (август 2000 г.). «Адаптация новых функций ферментов путем рациональной модернизации». Текущее мнение в структурной биологии. 10 (4): 405–10. Дои:10.1016 / S0959-440X (00) 00106-8. PMID 10981626.
- ^ Горг А., Вайс В., Данн М.Дж. (декабрь 2004 г.). «Современная технология двумерного электрофореза для протеомики». Протеомика. 4 (12): 3665–85. Дои:10.1002 / pmic.200401031. PMID 15543535. S2CID 28594824.
- ^ Конротто П., Сучельницкий С. (сентябрь 2008 г.). «Протеомные подходы в биологических и медицинских науках: принципы и приложения». Экспериментальная онкология. 30 (3): 171–80. PMID 18806738.
- ^ Koegl M, Uetz P (декабрь 2007 г.). «Улучшение двугибридных систем скрининга дрожжей». Брифинги по функциональной геномике и протеомике. 6 (4): 302–12. Дои:10.1093 / bfgp / elm035. PMID 18218650. В архиве из оригинала на 2017-09-11. Получено 2017-07-23.
- ^ Плевчинский Д., Гинальский К. (2009). «Интерактом: прогнозирование белок-белковых взаимодействий в клетках». Письма о клеточной и молекулярной биологии. 14 (1): 1–22. Дои:10.2478 / s11658-008-0024-7. ЧВК 6275871. PMID 18839074.
- ^ Чжан Ц., Ким Ш. (февраль 2003 г.). «Обзор структурной геномики: от структуры к функции». Современное мнение в области химической биологии. 7 (1): 28–32. Дои:10.1016 / S1367-5931 (02) 00015-7. PMID 12547423. В архиве из оригинала 2018-11-19. Получено 2019-06-29.
- ^ Бранден и Туз, стр. 340–41.
- ^ Гонен Т., Ченг Ю., Слиз П., Хироаки Ю., Фудзиёси И., Харрисон С.К., Уолз Т. (декабрь 2005 г.). «Липидно-белковые взаимодействия в двухслойных двумерных кристаллах AQP0». Природа. 438 (7068): 633–38. Bibcode:2005Натура.438..633Г. Дои:10.1038 / природа04321. ЧВК 1350984. PMID 16319884.
- ^ Standley DM, Kinjo AR, Kinoshita K, Nakamura H (июль 2008 г.). «Базы данных структуры белков с новыми веб-сервисами для структурной биологии и биомедицинских исследований». Брифинги по биоинформатике. 9 (4): 276–85. Дои:10.1093 / bib / bbn015. PMID 18430752. В архиве из оригинала 2013-04-15. Получено 2009-04-13.
- ^ Валиан П., Кросс Т.А., Япончик Б.К. (2004). «Структурная геномика мембранных белков». Геномная биология. 5 (4): 215. Дои:10.1186 / gb-2004-5-4-215. ЧВК 395774. PMID 15059248.
- ^ Слеатор Р.Д. (2012). «Прогнозирование функций белков». Функциональная геномика. Методы молекулярной биологии. 815. С. 15–24. Дои:10.1007/978-1-61779-424-7_2. ISBN 978-1-61779-423-0. PMID 22130980.
- ^ Чжан И (июнь 2008 г.). «Прогресс и проблемы в предсказании структуры белка». Текущее мнение в структурной биологии. 18 (3): 342–48. Дои:10.1016 / j.sbi.2008.02.004. ЧВК 2680823. PMID 18436442.
- ^ Сян Зи (июнь 2006 г.). «Достижения в моделировании структуры гомологичных белков». Современная наука о белках и пептидах. 7 (3): 217–27. Дои:10.2174/138920306777452312. ЧВК 1839925. PMID 16787261.
- ^ Чжан Ю., Сколник Дж. (Январь 2005 г.). «Проблема предсказания структуры белка может быть решена с использованием текущей библиотеки PDB». Труды Национальной академии наук Соединенных Штатов Америки. 102 (4): 1029–34. Bibcode:2005PNAS..102.1029Z. Дои:10.1073 / pnas.0407152101. ЧВК 545829. PMID 15653774.
- ^ Кульман Б., Дантас Г., Иретон Г.К., Варани Г., Стоддард Б.Л., Бейкер Д. (ноябрь 2003 г.). «Дизайн новой глобулярной белковой складки с точностью до атомного уровня». Наука. 302 (5649): 1364–68. Bibcode:2003Наука ... 302.1364K. Дои:10.1126 / science.1089427. PMID 14631033. S2CID 1939390.
- ^ Ричи DW (февраль 2008 г.). «Последние достижения и будущие направления в белок-белковой стыковке». Современная наука о белках и пептидах. 9 (1): 1–15. CiteSeerX 10.1.1.211.4946. Дои:10.2174/138920308783565741. PMID 18336319.
- ^ Щерага Х.А., Халили М, Ливо А (2007). «Динамика сворачивания белков: обзор методов молекулярного моделирования». Ежегодный обзор физической химии. 58: 57–83. Bibcode:2007ARPC ... 58 ... 57S. Дои:10.1146 / annurev.physchem.58.032806.104614. PMID 17034338.
- ^ Загрович Б., Snow CD, Shirts MR, Pande VS (ноябрь 2002 г.). «Моделирование сворачивания небольшого альфа-спирального белка в атомистических деталях с использованием всемирно распределенных вычислений». Журнал молекулярной биологии. 323 (5): 927–37. CiteSeerX 10.1.1.142.8664. Дои:10.1016 / S0022-2836 (02) 00997-X. PMID 12417204.
- ^ Хергес Т., Венцель В. (январь 2005 г.). «In silico сворачивание трехспирального белка и характеристика его ландшафта свободной энергии в полностью атомном силовом поле». Письма с физическими проверками. 94 (1): 018101. arXiv:физика / 0310146. Bibcode:2005PhRvL..94a8101H. Дои:10.1103 / PhysRevLett.94.018101. PMID 15698135. S2CID 1477100.
- ^ Hoffmann M, Wanko M, Strodel P, König PH, Frauenheim T., Schulten K, Thiel W, Tajkhorshid E, Elstner M (август 2006 г.). «Настройка цвета в родопсинах: механизм спектрального сдвига между бактериородопсином и сенсорным родопсином II». Журнал Американского химического общества. 128 (33): 10808–18. Дои:10.1021 / ja062082i. PMID 16910676.
- ^ Уорд Дж. Дж., Соди Дж. С., Макгаффин Л. Дж., Бакстон Б. Ф., Джонс Д. Т. (март 2004 г.). «Прогнозирование и функциональный анализ нативного расстройства в белках трех царств жизни». Журнал молекулярной биологии. 337 (3): 635–45. CiteSeerX 10.1.1.120.5605. Дои:10.1016 / j.jmb.2004.02.002. PMID 15019783.
- ^ Томпа П., Фершт А. (18 ноября 2009 г.). Структура и функция белков с внутренними нарушениями. CRC Press. ISBN 978-1-4200-7893-0. В архиве из оригинала 19 апреля 2017 г.. Получено 19 октября 2016.
- ^ Броснан Дж. Т. (июнь 2003 г.). «Межорганный транспорт аминокислот и его регуляция». Журнал питания. 133 (6 Прил. 1): 2068S – 72S. Дои:10.1093 / jn / 133.6.2068S. PMID 12771367.
- ^ Уотсон Т.Д. (1998). «Диета и кожные заболевания у собак и кошек». Журнал питания. 128 (12 Прил.): 2783S – 89S. Дои:10.1093 / jn / 128.12.2783S. PMID 9868266.
- ^ а б Case LP, Даристотель Л., Хайек М.Г., Рааш М.Ф. (2010). Электронная книга по питанию собак и кошек: ресурс для специалистов-домашних животных. Elsevier Health Sciences.
- ^ Muñoz-Huerta et al. (2013) Обзор методов определения статуса азота в растениях: преимущества, недостатки и последние достижения
- ^ Martin et al. (2002) Определение органического углерода и азота почвы на уровне поля с помощью спектроскопии в ближней инфракрасной области.
Учебники
- Бранден С., Туз Дж. (1999). Введение в структуру белка. Нью-Йорк: Garland Pub. ISBN 978-0-8153-2305-1.
- Мюррей РФ, Харпер Х.В., Граннер Д.К., Мэйс ПА, Родвелл Ф.В. (2006). Иллюстрированная биохимия Харпера. Нью-Йорк: Lange Medical Books / McGraw-Hill. ISBN 978-0-07-146197-9.
- Ван Холд К. Э., Мэтьюз К. К. (1996). Биохимия. Менло-Парк, Калифорния: Паб Бенджамин / Каммингс. Co., Inc. ISBN 978-0-8053-3931-4.
внешняя ссылка
Базы данных и проекты
- База данных NCBI Entrez Protein
- База данных NCBI Protein Structure
- Справочная база данных белков человека
- Человеческая протеинпедия
- Folding @ Home (Стэнфордский университет)
- Банк данных белков в Европе (смотрите также PDBeQuips, короткие статьи и руководства по интересным структурам PDB)
- Коллаборация исследований структурной биоинформатики (смотрите также Молекула месяца, представляя короткие отчеты по выбранным белкам из PDB)
- Протеопедия - жизнь в 3D: вращающаяся, масштабируемая 3D-модель с вики-аннотациями для каждой известной молекулярной структуры белка.
- UniProt - универсальный протеиновый ресурс
Учебники и образовательные сайты
- «Введение в белки» из НАДЕЖДЫ (Образовательный проект Хантингтона в области образования в Стэнфорде)
- Белки: биогенез до деградации - виртуальная библиотека биохимии и клеточной биологии
- Протеин на britannica.com
| Авторитетный контроль |
|---|