Кариотип - Karyotype
Кариотипирование это процесс, с помощью которого фотографии хромосомы берутся для определения хромосомного набора человека, включая количество хромосом и любые отклонения. Этот термин также используется для обозначения полного набора хромосом в виды или в отдельном организме[1][2][3] и для теста, который обнаруживает это дополнение или измеряет число.
Кариотипы описывают количество хромосом в организме и как эти хромосомы выглядят под светом микроскоп. Обращают внимание на их длину, положение центромеры, рисунок полос, любые различия между половые хромосомы, и любые другие физические характеристики.[4] Подготовка и исследование кариотипов является частью цитогенетика.
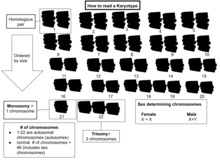

Изучение целых наборов хромосом иногда называют кариология. Хромосомы изображены (путем перестановки микрофотографии) в стандартном формате, известном как кариограмма или идиограмма: попарно, упорядочено по размеру и положению центромеры для хромосом одинакового размера.
Основное количество хромосом в соматический клетки человека или вида называется соматическое число и обозначен 2n. в зародышевый путь (половые клетки) номер хромосомы п (люди: n = 23).[2]стр.28 Таким образом, в люди 2n = 46.
Итак, в нормальном диплоид организмы аутосомный хромосомы присутствуют в двух копиях. Может быть, а может и не быть половые хромосомы. Полиплоид клетки имеют несколько копий хромосом и гаплоидный клетки имеют единичные экземпляры.
Кариотипы можно использовать для многих целей; например, чтобы учиться хромосомные аберрации, Сотовая связь функция таксономический отношения, лекарство и собрать информацию о прошлом эволюционный События (кариосистематика ).[5]
История изучения кариотипа
Хромосомы были впервые обнаружены в клетках растений Карл Вильгельм фон Нэгели в 1842 г. Их поведение у животных (саламандра ) клеток был описан Вальтер Флемминг, первооткрыватель митоз, в 1882 году. Название было придумано другим немецким анатомом, Генрих фон Вальдейер в 1888 году. Новая латынь от Древнегреческий κάρυον карион, «ядро», «семя» или «ядро» и τύπος опечатки, «общая форма»)
Следующий этап наступил после развития генетики в начале 20 века, когда стало понятно, что хромосомы (которые можно наблюдать по кариотипу) являются носителями генов. Лев Делоне в 1922 году, кажется, был первым, кто определил кариотип как фенотипический появление соматический хромосомы, в отличие от их генный содержание.[6][7] Последующую историю концепции можно проследить в работах К. Д. Дарлингтон[8] и Майкл Дж. Д. Уайт.[2][9]
Изучение кариотипа человека заняло много лет, чтобы решить самый главный вопрос: сколько хромосом делает нормальный диплоид клетка человека содержит?[10] В 1912 г. Ханс фон Винивартер сообщили о 47 хромосомах в сперматогония и 48 в оогония, завершая XX / XO определение пола механизм.[11] Художник в 1922 году не было уверенности, диплоид человека 46 или 48, сначала отдавая предпочтение 46,[12] но пересмотрел свое мнение с 46 на 48, и он правильно настаивал на том, что у людей есть XX / XY система.[13] Учитывая методы того времени, эти результаты были замечательными.
Джо Хин Тжио работает в Альберт Леван лаборатория[14] обнаружил, что количество хромосом составляет 46, используя новые доступные в то время методы:
- Использование ячеек в культура ткани
- Предварительная обработка клеток в гипотонический раствор, что увеличивает их и расширяет хромосомы
- Арест митоз в метафаза решением колхицин
- Сдавливание препарата на слайде, вынуждающее хромосомы в одну плоскость
- Разрезание микрофотографии и преобразование результата в бесспорную кариограмму.
Работа состоялась в 1955 году и была опубликована в 1956 году. Кариотип человека включает всего 46 хромосом.[15][16] Другой большие обезьяны имеют 48 хромосом. Хромосома человека 2 теперь известно, что это результат слияния двух предковых хромосом обезьян.[17][18]
Наблюдения за кариотипами
Окрашивание
Изучение кариотипов стало возможным благодаря окрашивание. Обычно подходящий красить, такие как Гимза,[19] применяется после клетки были арестованы во время деление клеток решением колхицин обычно в метафаза или прометафаза когда наиболее сжато. Для того, чтобы Гимза Чтобы краситель прилипал правильно, все хромосомные белки должны быть переварены и удалены. Для людей, белые кровяные клетки используются наиболее часто, потому что их легко заставить делиться и расти в культура ткани.[20] Иногда могут быть сделаны наблюдения по поводу неделимости (межфазный ) клетки. Пол нерожденного плод можно определить путем наблюдения за интерфазными ячейками (см. амниотический центез и Тело Барра ).
Наблюдения
Обычно наблюдают и сравнивают шесть различных характеристик кариотипов:[21]
- Различия в абсолютных размерах хромосом. Хромосомы могут различаться по абсолютному размеру до двадцати раз между родами одного и того же семейства. Например, бобовые Lotus tenuis и Vicia faba у каждого по шесть пар хромосом, но В. Фаба хромосомы во много раз больше. Эти различия, вероятно, отражают разную степень дупликации ДНК.
- Различия в положении центромеры. Эти различия, вероятно, возникли из-за транслокации.
- Различия в относительном размере хромосом. Эти различия, вероятно, возникли из-за сегментарного обмена неравной длины.
- Различия в основном количестве хромосом. Эти различия могли быть результатом последовательных неравных транслокаций, которые удаляли весь существенный генетический материал из хромосомы, допуская его потерю без ущерба для организма (гипотеза дислокации) или путем слияния. У людей на одну пару хромосом меньше, чем у человекообразных обезьян. Человеческая хромосома 2, по-видимому, возникла в результате слияния двух наследственных хромосом, и многие гены этих двух исходных хромосом были перемещены в другие хромосомы.
- Различия в количестве и положении спутников. Сателлиты - это маленькие тела, прикрепленные к хромосоме тонкой нитью.
- Различия в степени и распределении гетерохроматический регионы. Гетерохроматин окрашивается темнее, чем эухроматин. Гетерохроматин упакован плотнее. Гетерохроматин состоит в основном из генетически неактивных и повторяющихся последовательностей ДНК, а также содержит большее количество Аденин -Тимин пары. Эухроматин обычно находится в активной транскрипции и окрашивается намного светлее, так как он имеет меньшее сродство к Giemsa пятно.[22] Области эухроматина содержат большее количество Гуанин -Цитозин пары. Техника окрашивания с использованием Giemsa окрашивание называется G полосы и поэтому производит типичные "G-диапазоны".[22]
Таким образом, полное описание кариотипа может включать количество, тип, форму и полосатость хромосом, а также другую цитогенетическую информацию.
Часто встречаются вариации:
- между полами,
- между зародышевый путь и сома (между гаметы и остальное тело),
- между членами населения (полиморфизм хромосом ),
- в географическая специализация, и
- в мозаика или другие ненормальные люди.[9]
Кариотип человека

Типичные кариотипы человека содержат 22 пары аутосомный хромосомы и одна пара половые хромосомы (аллосомы). Наиболее распространенные кариотипы для самки содержать два Х-хромосомы и обозначены 46, XX; самцы обычно имеют как X, так и Y-хромосома обозначено 46, XY. Примерно 1,7% процентов людей интерсекс, иногда из-за различий в половых хромосомах.[23][24]
Некоторые вариации кариотипа, будь то аутосомы или аллосомы, вызывают аномалии развития.
Разнообразие и эволюция кариотипов
Хотя репликация и транскрипция из ДНК очень стандартизирован в эукариоты этого нельзя сказать об их кариотипах, которые сильно варьируют. Между видами существуют различия в количестве хромосом и в детальной организации, несмотря на то, что они построены из одного и того же макромолекулы. Эта вариация служит основой для ряда исследований в области эволюции. цитология.
В некоторых случаях даже между видами есть значительные различия. В своем обзоре Годфри и Мастерс заключают:
На наш взгляд, маловероятно, что тот или иной процесс может независимо объяснять широкий спектр наблюдаемых структур кариотипа ... Но в сочетании с другими филогенетическими данными, кариотипическое деление может помочь объяснить резкие различия в диплоидных числах. между близкородственными видами, что ранее было необъяснимо.[25]
Хотя многое известно о кариотипах на описательном уровне, и ясно, что изменения в организации кариотипов повлияли на эволюционный курс многих видов, совершенно неясно, какое может быть общее значение.
У нас очень плохое понимание причин эволюции кариотипа, несмотря на многочисленные тщательные исследования ... общее значение эволюции кариотипа неясно.
— Мэйнард Смит[26]
Изменения в процессе разработки
Вместо обычной репрессии генов некоторые организмы занимаются крупномасштабным устранением гетерохроматин или другие виды видимой адаптации к кариотипу.
- Удаление хромосом. У некоторых видов, как и у многих сциаридные мухи, целые хромосомы удаляются во время развития.[27]
- Уменьшение хроматина (отец-основатель: Теодор Бовери ). В этом процессе в некоторых копеподы и круглые черви такие как Ascaris suum части хромосом отбрасываются в определенных клетках. Этот процесс представляет собой тщательно организованную перестройку генома, при которой конструируются новые теломеры и теряются определенные участки гетерохроматина.[28][29] В А. суумвсе предшественники соматических клеток подвергаются уменьшению хроматина.[30]
- X-инактивация. Инактивация одной Х-хромосомы происходит во время раннего развития млекопитающих (см. Тело Барра и компенсация дозировки ). В плацентарные млекопитающие, инактивация случайна как между двумя X; таким образом, самка млекопитающего представляет собой мозаику в отношении ее Х-хромосом. В сумчатые всегда инактивирован отцовский X. У женщин около 15% соматических клеток избегают инактивации,[31] а количество генов, затронутых инактивированной Х-хромосомой, варьируется между клетками: в фибробласт клетки около 25% генов тела Барра избегают инактивации.[32]
Количество хромосом в наборе
Ярким примером изменчивости между близкородственными видами является мунтжак, который исследовал Курт Бениршке и Дорис Вурстер. Диплоидное число китайского мунтжака, Muntiacus reevesi, оказалось 46, все телоцентрический. Когда они посмотрели на кариотип близкого индийского мунтжака, Muntiacus muntjak, они были удивлены, обнаружив, что у него женские = 6, мужские = 7 хромосом.[33]
Они просто не могли поверить в то, что видели ... Они молчали два или три года, потому что думали, что что-то не так с их тканевой культурой ... Но когда они получили еще пару образцов, они подтвердили [свои выводы].
— Hsu p. 73-4[16]
Число хромосом в кариотипе между (относительно) неродственными видами сильно варьируется. Самый низкий рекорд принадлежит нематода Parascaris univalens, где гаплоидный п = 1; и муравей: Myrmecia pilosula.[34] Высокий рекорд будет где-то среди папоротники, папоротник с языком гадюки Офиоглоссум впереди в среднем 1262 хромосомы.[35] Наивысший балл для животных может быть коротконосый осетр Acipenser brevirostrum на 372 хромосомах.[36] Наличие нештатных или В-хромосомы означает, что число хромосом может варьироваться даже в пределах одной межпородной популяции; и анеуплоиды являются другим примером, хотя в этом случае они не будут рассматриваться как нормальные члены населения.
Основное число
Основное число, FN, кариотипа - это количество видимых основных хромосомных плеч на набор хромосом.[37][38] Таким образом, FN ≤ 2 x 2n, различие зависит от количества хромосом, считающихся одноплечими (акроцентрический или телоцентрический ) подарок. У людей FN = 82,[39] из-за наличия пяти пар акроцентрических хромосом: 13, 14, 15, 21, и 22 (человек Y-хромосома также акроцентрический). Основное аутосомное число или основное аутосомное число, FNa[40] или AN,[41] кариотипа - это количество видимых основных хромосомных плеч на набор аутосомы (несцепленные с полом хромосомы ).
Плоидность
Плоидность количество полных наборов хромосом в клетке.
- Полиплоидия, где в клетках имеется более двух наборов гомологичных хромосом, встречается в основном у растений. По мнению авторов, он имел большое значение в эволюции растений. Стеббинс.[42][43][44][45] Доля цветковых растений, которые являются полиплоидными, по оценке Стеббинса, составляет 30–35%, но у злаков средний показатель намного выше, около 70%.[46] Полиплоидия низших растений (папоротники, хвощи и псилоталы ) также является обычным явлением, и некоторые виды папоротников достигли уровней полиплоидии, намного превышающих самые высокие уровни, известные для цветковых растений.
Полиплоидия у животных встречается гораздо реже, но в некоторых группах она значительна.[47]
Полиплоидные серии у родственных видов, которые полностью состоят из кратных единственного основного числа, известны как эуплоид.
- Гапло-диплоидия, где один пол диплоид, и другие гаплоидный. Это обычная договоренность в Перепончатокрылые, и в некоторых других группах.
- Эндополиплоидия возникает, когда у взрослого дифференцированный ткани, на которые клетки перестали делиться митоз, но ядра содержат больше, чем оригинал соматический количество хромосомы.[48] в эндоцикл (эндомитоз или эндоредупликация ) хромосомы в "покоящемся" ядре подвергаются дублирование, дочерние хромосомы, отделяющиеся друг от друга внутри неповрежденный ядерная мембрана.[49]
Во многих случаях эндополиплоидные ядра содержат десятки тысяч хромосом (которые невозможно точно подсчитать). Клетки не всегда содержат точные кратные (степени двойки), поэтому простое определение «увеличение числа наборов хромосом, вызванное репликацией без деления клетки» не совсем точное.
Этот процесс (особенно изученный у насекомых и некоторых высших растений, таких как кукуруза) может быть стратегией развития для увеличения продуктивности тканей, которые очень активны в биосинтезе.[50]
Это явление спорадически возникает на всем протяжении эукариот королевство из простейшие людям; он разнообразен и сложен и служит дифференциация и морфогенез во многих отношениях.[51] - Увидеть палеополиплоидия для исследования дупликаций древних кариотипов.
Анеуплоидия
Анеуплоидия это состояние, при котором количество хромосом в клетках не является типичным для данного вида. Это привело бы к хромосомная аномалия например, лишняя хромосома или потеря одной или нескольких хромосом. Аномалии числа хромосом обычно вызывают дефект в развитии. Синдром Дауна и Синдром Тернера примеры этого.
Анеуплоидия также может возникать в группе близкородственных видов. Классическими примерами у растений являются род Crepis, где гаметические (= гаплоидные) числа образуют ряды x = 3, 4, 5, 6 и 7; и Крокус, где каждое число от x = 3 до x = 15 представлено как минимум одним видом. Разного рода свидетельства показывают, что тенденции эволюции в разных группах разошлись в разных направлениях.[52] У приматов большие обезьяны имеют хромосомы 24x2, тогда как у людей 23x2. Хромосома человека 2 был образован в результате слияния наследственных хромосом, уменьшив количество.[53]
Хромосомный полиморфизм
Некоторые виды полиморфный для разных структурных форм хромосом.[54] Структурная изменчивость может быть связана с разным количеством хромосом у разных людей, что встречается у жуков-божьих коровок. Клеймо хилокоруса, немного богомолы рода Амелес,[нужна цитата ] европейская землеройка Sorex araneus.[55] Есть некоторые свидетельства из дела моллюск Тайский лапиллус (в собачий прыщ ) на Бретань Коста, что две морфы хромосом адаптированный в разные места обитания.[56]
Видовые деревья
Детальное изучение полос хромосом у насекомых с политенные хромосомы может выявить отношения между близкородственными видами: классическим примером является изучение полос хромосом на Гавайях. дрозофилиды от Хэмптон Л. Карсон.
Примерно через 17000 км2), Гавайские острова имеют самую разнообразную в мире коллекцию мух-дрозофилид, живущих с тропические леса к субальпийские луга. Эти примерно 800 гавайских видов дрозофилид обычно относятся к двум родам: Дрозофила и Скаптомиза, в семье Drosophilidae.
Политеновая полосатость группы «картинное крыло», наиболее изученной группы гавайских дрозофилид, позволила Карсону построить эволюционное древо задолго до того, как стал возможен анализ генома. В некотором смысле расположение генов можно увидеть в полосах каждой хромосомы. Хромосомные перестройки, особенно инверсии, позволяют увидеть, какие виды тесно связаны между собой.
Результаты очевидны. Инверсии, когда они построены в виде дерева (и не зависят от всей другой информации), показывают четкий «поток» видов от старых островов к новым. Бывают также случаи возврата к более старым островам и обхода островов, но они гораздо реже. С помощью K-Ar датировка, нынешние острова датируются 0,4 миллиона лет назад (млн лет назад) (Мауна-Кеа ) до 10мя (Неккер ). Самый старый член Гавайского архипелага, все еще возвышающийся над морем, Атолл Куре, который можно датировать 30 млн лет назад. Сам архипелаг (произведен Тихоокеанская плита переезд горячая точка ) существует гораздо дольше, по крайней мере, в Меловой. Предыдущие острова теперь под морем (гайоты ) образуют Императорская цепь подводных гор.[57]
Все родное Дрозофила и Скаптомиза Виды на Гавайях, по-видимому, произошли от единственного предкового вида, колонизировавшего острова, вероятно, 20 миллионов лет назад. Последующие адаптивное излучение был подстегнут отсутствием соревнование и широкий выбор ниши. Хотя можно было бы и сингл беременный Самка колонизировала остров, скорее всего, это была группа одного и того же вида.[58][59][60][61]
На Гавайском архипелаге есть и другие животные и растения, которые подверглись подобным, хотя и менее впечатляющим, адаптивным излучениям.[62][63]
Хромосомные полосы
Хромосомы демонстрируют полосатый рисунок при обработке некоторых пятен. Полосы - это чередующиеся светлые и темные полосы, которые появляются по длине хромосом. Уникальные образцы полос используются для идентификации хромосом и диагностики хромосомных аберраций, включая разрыв хромосомы, потерю, дупликацию, транслокацию или инвертированные сегменты. Целый ряд различных хромосомных обработок приводит к появлению ряда паттернов полос: G-диапазоны, R-диапазоны, C-диапазоны, Q-диапазоны, T-диапазоны и NOR-диапазоны.
Изображение кариотипов
Виды бандажа
Цитогенетика использует несколько методов для визуализации различных аспектов хромосом:[20]
- G-полосы получается с Пятно Гимзы после переваривания хромосом трипсин. Он дает серию светлых и темных полос - темные области, как правило, гетерохроматичны, реплицируются поздно и богаты АТ. Светлые области имеют тенденцию быть эухроматическими, рано реплицирующимися и богатыми GC. Этот метод обычно дает 300–400 полос в нормальном, человеческий геном.
- R-banding - это обратное G-banding (R означает «обратный»). Темные области являются эухроматическими (области, богатые гуанином-цитозином), а светлые области - гетерохроматическими (области, богатые тимин-аденином).
- C-banding: Giemsa связывается с конститутивный гетерохроматин, так он пачкает центромеры. Название происходит от центромерного или конститутивного гетерохроматина. Перед окрашиванием препараты подвергаются щелочной денатурации, что приводит к почти полной депуринизации ДНК. После промывки зонда оставшуюся ДНК снова ренатурируют и окрашивают раствором Гимза, состоящим из метиленазура, метиленового фиолетового, метиленового синего и эозина. Гетерохроматин связывает много красителя, тогда как остальные хромосомы поглощают его лишь немного. С-связывание оказалось особенно подходящим для характеристики хромосом растений.
- Q-бэндинг - это флуоресцентный образец получен с использованием хинакрин для окрашивания. Структура полос очень похожа на полосу G. Их можно распознать по желтой флуоресценции разной интенсивности. Большая часть окрашенной ДНК - гетерохроматин. Хинакрин (атебрин) связывает обе области, богатые AT и GC, но флуоресцирует только комплекс AT-хинакрина. Поскольку области, богатые AT, более обычны в гетерохроматине, чем в эухроматине, эти области предпочтительно помечены. Различная интенсивность отдельных полос отражает различное содержание AT. Другие флуорохромы, такие как DAPI или Hoechst 33258, также дают характерные воспроизводимые образцы. Каждый из них дает свой особый узор. Другими словами: свойства связей и специфичность флуорохромов не основываются исключительно на их сродстве к областям, богатым AT. Напротив, распределение AT и ассоциация AT с другими молекулами, такими как, например, гистоны, влияет на связывающие свойства флуорохромов.
- Т-образные полосы: визуализировать теломеры.
- Окрашивание серебром: Нитрат серебра окрашивает область ядрышковой организации -ассоциированный белок. Это дает темную область, где осаждается серебро, что указывает на активность генов рРНК в ЯОР.
Классическая цитогенетика кариотипа

В «классическом» (изображенном) кариотипе a красить, довольно часто Гимза (G-полосы), менее часто мепакрин (хинакрин), используется для окрашивания полос на хромосомах. Гимза специфичен для фосфат группы ДНК. Хинакрин связывается с аденин -тимин -богатые регионы. Каждая хромосома имеет характерный рисунок полос, который помогает их идентифицировать; обе хромосомы в паре будут иметь одинаковый рисунок полос.
Кариотипы расположены так, что короткое плечо хромосомы находится наверху, а длинное плечо - внизу. Некоторые кариотипы называют короткие и длинные руки п и qсоответственно. Кроме того, различным окрашенным областям и подобластям даются числовые обозначения от проксимальный к дистальный на плечах хромосом. Например, Кри дю чат синдром включает делецию на коротком плече хромосомы 5. Он записывается как 46, XX, 5p-. Критическая область для этого синдрома - делеция p15.2 ( локус на хромосоме), который записывается как 46, XX, del (5) (p15.2).[64]
Многоцветный FISH (mFISH) и спектральный кариотип (метод SKY)
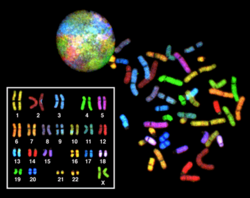
Многоцветный РЫБЫ и более старое спектральное кариотипирование является молекулярным цитогенетический методы, используемые для одновременной визуализации всех пар хромосомы в организме разными цветами. Флуоресцентно меченые пробы для каждой хромосомы изготавливаются путем маркировки хромосомно-специфической ДНК разными флуорофоры. Поскольку существует ограниченное количество спектрально различных флуорофоров, для получения множества различных цветов используется комбинаторный метод маркировки. Комбинации флуорофоров фиксируются и анализируются с помощью флуоресцентного микроскопа с использованием до 7 узкополосных флуоресцентных фильтров или, в случае спектрального кариотипирования, с использованием интерферометр прикреплен к флуоресцентному микроскопу. В случае изображения mFISH каждая комбинация флуорохромов из полученных исходных изображений заменяется псевдоцвет в специальном программном обеспечении для анализа изображений. Таким образом, можно визуализировать и идентифицировать хромосомы или участки хромосом, что позволяет анализировать хромосомные перестройки.[65]В случае спектрального кариотипирования программа обработки изображений назначает псевдоцвет для каждой спектрально разной комбинации, что позволяет визуализировать индивидуально окрашенные хромосомы.[66]

Многоцветный FISH используется для определения структурных хромосомных аберраций в раковых клетках и других болезненных состояний, когда полосатость Гимзы или другие методы недостаточно точны.
Цифровое кариотипирование
Цифровое кариотипирование это метод, используемый для количественной оценки количества копий ДНК в геномной шкале. Выделяют и перечисляют короткие последовательности ДНК из определенных локусов по всему геному.[67] Этот метод также известен как виртуальное кариотипирование.
Хромосомные аномалии
Хромосомные аномалии могут быть числовыми, как в случае наличия лишних или отсутствующих хромосом, или структурными, как в производная хромосома, транслокации, инверсии, крупномасштабные удаления или дублирования. Числовые отклонения, также известные как анеуплоидия, часто возникают в результате нерасхождение в течение мейоз в формировании гамета; трисомии, в которых присутствуют три копии хромосомы вместо обычных двух, являются обычными числовыми аномалиями. Структурные аномалии часто возникают из-за ошибок в гомологичная рекомбинация. Оба типа аномалий могут возникать в гаметах и, следовательно, будут присутствовать во всех клетках тела пострадавшего человека, или они могут возникать во время митоз и дать начало генетическая мозаика человек, у которого есть нормальные и некоторые аномальные клетки.
В людях
Хромосомные аномалии, которые приводят к заболеваниям у людей, включают:
- Синдром Тернера является результатом одной X-хромосомы (45, X или 45, X0).
- Синдром Клайнфельтера, наиболее распространенное мужское хромосомное заболевание, известное как 47, XXY, вызывается дополнительным Икс хромосома.
- Синдром Эдвардса вызвано трисомия (три копии) хромосомы 18.
- Синдром Дауна, распространенное хромосомное заболевание, вызывается трисомией 21 хромосомы.
- Синдром Патау вызвано трисомией хромосомы 13.
- Трисомия 9, считается 4-й по распространенности трисомией, у многих долгожителей поражены люди, но только в форме, отличной от полной трисомии, такой как синдром трисомии 9p или мозаичная трисомия 9. Они часто функционируют довольно хорошо, но, как правило, имеют проблемы с речью. .
- Также задокументированы трисомия 8 и трисомия 16, хотя обычно они не доживают до рождения.
Некоторые нарушения возникают в результате потери только части одной хромосомы, в том числе
- Кри дю чат (крик кошки) от усеченной короткой руки на хромосоме 5.Название происходит от характерного крика младенцев, вызванного аномальным формированием гортани.
- 1p36 Синдром делеции, от потери части короткого плеча хромосомы 1.
- Синдром ангельмана - в 50% случаев отсутствует сегмент длинного плеча 15 хромосомы; делеция материнских генов, пример печать беспорядок.
- Синдром Прадера-Вилли - в 50% случаев отсутствует сегмент длинного плеча 15 хромосомы; делеция отцовских генов, пример импринтингового расстройства.
- Хромосомные аномалии также могут возникать в раковый клетки генетически нормального человека; один хорошо задокументированный пример - это Филадельфийская хромосома, транслокационная мутация, обычно связанная с хронический миелолейкоз и реже с острый лимфобластный лейкоз.
Смотрите также
использованная литература
- ^ Краткий оксфордский словарь
- ^ а б c Белый 1973, п. 35 год
- ^ Стеббинс, Г.Л. (1950). «Глава XII: Кариотип». Вариации и эволюция растений. Издательство Колумбийского университета.
- ^ King, R.C .; Стэнсфилд, W.D .; Маллиган, П. (2006). Словарь генетики (7-е изд.). Издательство Оксфордского университета. п.242.
- ^ «Кариосистематика».
- ^ Делоне Л. Н. Сравнительное кариологическое исследование видов Muscari Mill. и Беллевалия Лапейр. Вестник Тифлисского ботанического сада. 1922, т. 2, н. 1, стр. 1-32 [на русском]
- ^ Батталья, Эмилио (1994). «Нуклеосома и нуклеотип: терминологическая критика». Кариология. 47 (3–4): 193–197. Дои:10.1080/00087114.1994.10797297.
- ^ Дарлингтон С.Д. 1939 г. Эволюция генетических систем. Издательство Кембриджского университета. 2-е изд., Переработанное и дополненное, 1958 г. Оливер и Бойд, Эдинбург.
- ^ а б Белый M.J.D. 1973 г. Цитология и эволюция животных. 3-е изд., Cambridge University Press.
- ^ Коттлер MJ (1974). «От 48 до 46: цитологический метод, предубеждение и подсчет хромосом человека». Булл Хист Мед. 48 (4): 465–502. PMID 4618149.
- ^ фон Винивартер Х. (1912). "Études sur la spermatogenèse humaine". Archives de Biologie. 27 (93): 147–9.
- ^ Художник Т.С. (1922). «Сперматогенез человека». Анат. Res. 23: 129.
- ^ Художник Т.С. (1923). «Исследования сперматогенеза у млекопитающих II». J. Exp. Зоология. 37 (3): 291–336. Дои:10.1002 / jez.1400370303.
- ^ Райт, Пирс (11 декабря 2001 г.). "Джо Хин Тжио Человек, который вычислил количество хромосом". Хранитель.
- ^ Tjio J.H .; Леван А. (1956). «Число хромосом человека». Наследие. 42 (1–2): 1–6. Дои:10.1111 / j.1601-5223.1956.tb03010.x. PMID 345813.
- ^ а б Hsu T.C. 1979 г. Цитогенетика человека и млекопитающих: историческая перспектива. Спрингер-Верлаг, штат Нью-Йорк.
- ^ Хромосома 2 человека представляет собой слияние двух предков. хромосомы Алек МакЭндрю; по состоянию на 18 мая 2006 г.
- ^ Доказательства общего происхождения: хромосома 2 человека (видео) 2007
- ^ Препарат, в состав которого входят красители метиленовый синий, эозин Y и лазурь-A, B, C
- ^ а б Густашоу К.М. 1991. Хромосомные пятна. В Лабораторное руководство по цитогенетике ACT 2-е изд., Изд. М.Дж. Барч. Ассоциация цитогенетических технологов, Raven Press, Нью-Йорк.
- ^ Стеббинс, Г.Л. (1971). Хромосомная эволюция у высших растений. Лондон: Арнольд. стр.85–86.
- ^ а б Томпсон и Томпсон Генетика в медицине, 7-е изд.
- ^ "Насколько распространен интерсекс?". Кампания интерсексуалов за равенство. Получено 10 мая 2018.
- ^ «О количестве интерсекс людей». Intersex Human Rights Australia. Получено 10 мая 2018.
- ^ Годфри Л. Р., Мастерс Дж. С. (август 2000 г.). «Теория воспроизводства кинетохор может объяснить быструю эволюцию хромосом». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 97 (18): 9821–3. Bibcode:2000PNAS ... 97.9821G. Дои:10.1073 / пнас.97.18.9821. ЧВК 34032. PMID 10963652.
- ^ Мейнард Смит Дж. 1998. Эволюционная генетика. 2-е изд, Оксфорд. p218-9
- ^ Годай С., Эстебан М.Р. (март 2001 г.). «Удаление хромосом у сциаридных мух». BioEssays. 23 (3): 242–50. Дои:10.1002 / 1521-1878 (200103) 23: 3 <242 :: AID-BIES1034> 3.0.CO; 2-P. PMID 11223881.
- ^ Мюллер Ф, Бернар В., Тоблер Х (февраль 1996 г.). «Уменьшение хроматина у нематод». BioEssays. 18 (2): 133–8. Дои:10.1002 / bies.950180209. PMID 8851046. S2CID 24583845.
- ^ Wyngaard GA, Грегори TR (декабрь 2001 г.). «Временной контроль репликации ДНК и адаптивное значение уменьшения хроматина у копепод». J. Exp. Zool. 291 (4): 310–6. Дои:10.1002 / jez.1131. PMID 11754011.
- ^ Гилберт С.Ф. 2006 г. Биология развития. Sinauer Associates, Стэмфорд, штат Коннектикут. 8-е изд, Глава 9
- ^ Кинг, Стэнсфилд и Маллиган, 2006 г.
- ^ Каррел Л., Уиллард Х (2005). «Профиль X-инактивации показывает широкую вариабельность экспрессии X-сцепленного гена у женщин». Природа. 434 (7031): 400–404. Bibcode:2005 Натур. 434..400C. Дои:10.1038 / природа03479. PMID 15772666. S2CID 4358447.
- ^ Вурстер Д.Х., Бениршке К. (июнь 1970 г.). "Индийский мунтжак, Muntiacus muntjak: олень с низким диплоидным числом хромосом ». Наука. 168 (3937): 1364–6. Bibcode:1970Sci ... 168.1364W. Дои:10.1126 / science.168.3937.1364. PMID 5444269. S2CID 45371297.
- ^ Crosland M.W.J .; Крозье, Р. Х. (1986). "Myrmecia pilosula, муравей только с одной парой хромосом ». Наука. 231 (4743): 1278. Bibcode:1986Sci ... 231.1278C. Дои:10.1126 / science.231.4743.1278. PMID 17839565. S2CID 25465053.
- ^ Хандельвал С. (1990). «Хромосомная эволюция рода Ophioglossum L». Ботанический журнал Линнеевского общества. 102 (3): 205–217. Дои:10.1111 / j.1095-8339.1990.tb01876.x.
- ^ Kim, D.S .; Nam, Y.K .; Но, J.K .; Park, C.H .; Чепмен, Ф.А. (2005). "Кариотип североамериканского коротконосого осетра" Acipenser brevirostrum с наибольшим числом хромосом среди Acipenseriformes ". Ихтиологические исследования. 52 (1): 94–97. Дои:10.1007 / s10228-004-0257-z. S2CID 20126376.
- ^ Мэтти, Р. (15 мая 1945 г.). "L'evolution de la formule chromosomiale chez les vertébrés". Experientia (Базель). 1 (2): 50–56. Дои:10.1007 / BF02153623. S2CID 38524594.
- ^ de Oliveira, R.R .; Feldberg, E .; дос Аньос, М. Б.; Зуанон, Дж. (Июль – сентябрь 2007 г.). "Характеристика кариотипа и гетероморфизм половых хромосом ZZ / ZW у двух видов рода сомов" Анциструс Kner, 1854 (Siluriformes: Loricariidae) из бассейна Амазонки ». Неотропическая ихтиология. 5 (3): 301–6. Дои:10.1590 / S1679-62252007000300010.
- ^ Pellicciari, C .; Formenti, D .; Redi, C.A .; Manfredi, M.G .; Романини (февраль 1982 г.). «Изменчивость содержания ДНК у приматов». Журнал эволюции человека. 11 (2): 131–141. Дои:10.1016 / S0047-2484 (82) 80045-6.
- ^ Souza, A.L.G .; de O. Corrêa, M.M .; de Aguilar, C.T .; Пессоа, Л.М. (февраль 2011 г.). "Новый кариотип Wiedomys pyrrhorhinus (Rodentia: Sigmodontinae) из Чапада Диамантина, северо-восток Бразилии " (PDF). Зоология. 28 (1): 92–96. Дои:10.1590 / S1984-46702011000100013.
- ^ Weksler, M .; Бонвичино, C.R. (3 января 2005 г.). "Таксономия рода карликовых рисовых крыс Олигоризомис Bangs, 1900 (Rodentia, Sigmodontinae) бразильского Серрадо, с описанием двух новых видов » (PDF). Arquivos do Museu Nacional, Рио-де-Жанейро. 63 (1): 113–130. ISSN 0365-4508. Архивировано из оригинал (PDF) 26 марта 2012 г.. Получено 22 апреля 2012.
- ^ Стеббинс, Г.Л. (1940). «Значение полиплоидии в эволюции растений». Американский натуралист. 74 (750): 54–66. Дои:10.1086/280872. S2CID 86709379.
- ^ Стеббинс 1950
- ^ Comai L (ноябрь 2005 г.). «Преимущества и недостатки полиплоидности». Nat. Преподобный Жене. 6 (11): 836–46. Дои:10.1038 / nrg1711. PMID 16304599. S2CID 3329282.
- ^ Адамс К.Л., Вендель Дж.Ф. (апрель 2005 г.). «Полиплоидия и эволюция генома у растений». Curr. Мнение. Биол растений. 8 (2): 135–41. Дои:10.1016 / j.pbi.2005.01.001. PMID 15752992.
- ^ Стеббинс 1971
- ^ Gregory, T.R .; Мейбл, Б. (2011). «Глава 8: Полиплоидия у животных». В Грегори, Т. Райан (ред.). Эволюция генома. Академическая пресса. С. 427–517. ISBN 978-0-08-047052-8.
- ^ Уайт, M.J.D. (1973). Хромосомы (6-е изд.). Лондон: Чепмен и Холл. п.45.
- ^ Lilly M.A .; Дуронио Р.Дж. (2005). "Новое понимание контроля клеточного цикла из Дрозофила эндоцикл ". Онкоген. 24 (17): 2765–75. Дои:10.1038 / sj.onc.1208610. PMID 15838513.
- ^ Эдгар Б.А., Орр-Уивер Т.Л. (май 2001 г.). «Циклы эндорепликационных клеток: больше за меньшие деньги». Ячейка. 105 (3): 297–306. Дои:10.1016 / S0092-8674 (01) 00334-8. PMID 11348589. S2CID 14368177.
- ^ Нагл В. 1978. Эндополиплоидия и политения в дифференцировке и эволюции: к пониманию количественной и качественной изменчивости ядерной ДНК в онтогенезе и филогенезе. Эльзевир, Нью-Йорк.
- ^ Стеббинс, Дж. Ледли-младший, 1972 г. Хромосомная эволюция у высших растений. Нельсон, Лондон. p18
- ^ Айдо Дж. У., Бальдини А., Уорд, округ Колумбия, Ридерс СТ, Уэллс, РА (октябрь 1991 г.). "Происхождение хромосомы 2 человека: исконное слияние теломер и теломер". Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 88 (20): 9051–5. Bibcode:1991PNAS ... 88.9051I. Дои:10.1073 / пнас.88.20.9051. ЧВК 52649. PMID 1924367.
- ^ Rieger, R .; Michaelis, A .; Грин, М. (1968). Глоссарий генетики и цитогенетики: классическая и молекулярная. Нью-Йорк: Springer-Verlag. ISBN 9780387076683.
- ^ Сирл, Дж. Б. (1 июня 1984 г.). «Три новых кариотипических расы обыкновенной бурозубки Sorex Araneus (Mammalia: Insectivora) и филогения». Систематическая биология. 33 (2): 184–194. Дои:10.1093 / sysbio / 33.2.184. ISSN 1063-5157.
- ^ Белый 1973, п. 169
- ^ Clague, D.A .; Далримпл, Г. (1987). "Гавайско-Императорская вулканическая цепь. Часть I. Геологическая эволюция" (PDF). В Decker, R.W .; Wright, T.L .; Штауффер, П. (ред.). Вулканизм на Гавайях. 1. С. 5–54. Документ профессионала геологической службы США 1350.
- ^ Карсон Х.Л. (июнь 1970 г.). «Хромосомные индикаторы происхождения видов». Наука. 168 (3938): 1414–8. Bibcode:1970Sci ... 168.1414C. Дои:10.1126 / science.168.3938.1414. PMID 5445927.
- ^ Карсон Х.Л. (март 1983 г.). "Хромосомные последовательности и межостровная колонизация на Гавайях. Дрозофила". Генетика. 103 (3): 465–82. ЧВК 1202034. PMID 17246115.
- ^ Карсон Х.Л. (1992). "Инверсии на гавайском Дрозофила". In Krimbas, C.B .; Powell, J.R. (ред.). Полиморфизм инверсии дрозофилы. Бока-Ратон, Флорида: CRC Press. С. 407–439. ISBN 978-0849365478.
- ^ Канеширо, К.Ю .; Gillespie, R.G .; Карсон, Х.Л. (1995). «Хромосомы и мужские гениталии гавайской дрозофилы: инструменты для интерпретации филогении и географии». В Wagner, W.L .; Функ, Э. (ред.). Гавайская биогеография: эволюция на архипелаге горячей точки. Вашингтон, округ Колумбия: Пресса Смитсоновского института. стр.57–71.
- ^ Крэддок Э.М. (2000). «Процессы видообразования в адаптивной радиации гавайских растений и животных». Эволюционная биология. 31. С. 1–43. Дои:10.1007/978-1-4615-4185-1_1. ISBN 978-1-4613-6877-9. Отсутствует или пусто
| название =(Помогите) - ^ Зиглер, Алан К. (2002). Гавайская естественная история, экология и эволюция. Гавайский университет Press. ISBN 978-0-8248-2190-6.
- ^ Лиза Г. Шаффер; Нильс Томмеруп, ред. (2005). ISCN 2005: Международная система цитогенетической номенклатуры человека. Швейцария: S. Karger AG. ISBN 978-3-8055-8019-9.
- ^ Liehr T., Starke H, Weise A, Lehrer H, Claussen U (январь 2004 г.). «Наборы многоцветных щупов FISH и их применение». Histol. Гистопатол. 19 (1): 229–237. PMID 14702191.
- ^ Шрек Э., дю Мануар С., Вельдман Т. и др. (Июль 1996 г.). «Многоцветное спектральное кариотипирование хромосом человека». Наука. 273 (5274): 494–7. Bibcode:1996Sci ... 273..494S. Дои:10.1126 / science.273.5274.494. PMID 8662537. S2CID 22654725.
- ^ Ван Т.Л., Майерхофер С., Спайхер М.Р. и др. (Декабрь 2002 г.). «Цифровое кариотипирование». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 99 (25): 16156–61. Bibcode:2002PNAS ... 9916156W. Дои:10.1073 / pnas.202610899. ЧВК 138581. PMID 12461184.
внешние ссылки
 СМИ, связанные с Кариотипы в Wikimedia Commons
СМИ, связанные с Кариотипы в Wikimedia Commons- Создание кариотипа, интерактивное мероприятие Центра изучения генетики Университета штата Юта.
- Кариотипирование с историями болезни из Биологического проекта Университета Аризоны.
- Проект кариотипа для печати из Biology Corner, ресурсного сайта для учителей биологии и естественных наук.
- Методы окрашивания и окантовки хромосом
- Биосистемы Бьорна для кариотипирования и FISH