Миоцен - Miocene
| Неоген | |
|---|---|
| 23.03 – 5.333 Ма | |
| Хронология | |
Ключевые события неогена -24 — – -22 — – -20 — – -18 — – -16 — – -14 — – -12 — – -10 — – -8 — – -6 — – -4 — – -2 — Приблизительная шкала времени ключевых событий неогена. Вертикальная ось: миллионы лет назад. | |
| Этимология | |
| Формальность имени | Формальный |
| Информация об использовании | |
| Небесное тело | земной шар |
| использование | Глобальный (ICS ) |
| Используемая шкала времени | Шкала времени ICS |
| Определение | |
| Хронологическая единица | Эпоха |
| Стратиграфическая единица | Серии |
| Формальность промежутка времени | Формальный |
| Определение нижней границы |
|
| Нижняя граница ГССП | Секция Лемме-Каррозио, Карросио, Италия 44 ° 39′32 ″ с.ш. 8 ° 50′11 ″ в.д. / 44,6589 ° с. Ш. 8,8364 ° в. |
| ГССП ратифицирован | 1996 |
| Определение верхней границы | База магнитного события Thvera (C3n.4n), которое всего на 96 тыс. Лет назад (5 циклов прецессии) моложе GSSP. |
| Верхняя граница ГССП | Секция Гераклея Миноа, Гераклея Миноа, Каттолика Эраклея, Сицилия, Италия 37 ° 23′30 ″ с.ш. 13 ° 16′50 ″ в.д. / 37,3917 ° с. Ш. 13,2806 ° в. |
| ГССП ратифицирован | 2000[3] |
В Миоцен (/ˈмаɪ.əˌsяп,ˈмаɪ.oʊ-/ МОЙ--видел, МОЙ-ой-[4][5]) является первым геологическая эпоха из Неоген Период и длится примерно с 23.03 - 5.333 миллион лет назад (Ма). Миоцен был назван Чарльз Лайель; его название происходит от греческих слов μείων (Meiōn, "меньше") и καινός (кайнос, "новый")[6][7] и означает «менее свежий», потому что в нем на 18% меньше современных морских беспозвоночные чем Плиоцен.[8] Миоцену предшествует Олигоцен и следует Плиоцен.
По мере того, как Земля перешла от олигоцена через миоцен к плиоцену, климат медленно охладился до ряда ледниковые периоды. Границы миоцена не отмечены каким-то одним отчетливым глобальным событием, а состоят скорее из региональных границ между более теплым олигоценом и более холодной эпохой плиоцена.
В обезьяны впервые возникла, возникла и разнообразилась во время ранний миоцен (Аквитанский и бурдигалский этапы), получившие широкое распространение в Старый мир. К концу этой эпохи и началу следующей эпохи предки людей откололись от предков шимпанзе следовать своему собственному эволюционному пути на заключительном мессинском этапе (7,5–5,3 млн лет) миоцена. Как и в предшествующий олигоцен, луга продолжали расширяться, а леса сокращаться. В морях миоцена, ламинарии леса впервые появились и вскоре стали одной из самых продуктивных экосистем Земли.[9]
Растения и животные миоцена были узнаваемо современными. Млекопитающие и птицы были хорошо известны. Киты, ластоногие, и ламинария распространять.
Миоцен представляет особый интерес для геологов и палеоклиматологов как основная фаза геология Гималаев произошел в миоцене, затронув муссонный узоры в Азии, которые были связаны с ледниковые периоды в северном полушарии.[10]
Подразделения

Миоцен фаунистические этапы от младшего к старшему обычно называют в соответствии с Международная комиссия по стратиграфии:[11]
| Субэпоха | Фаунистический этап | Временной диапазон |
|---|---|---|
| Поздний миоцен | Мессинианский | 7.246–5.333 Ма |
| Тортониан | 11,608–7,246 млн лет | |
| Средний миоцен | Серравальский | 13.65–11.608 млн лет |
| Langhian | 15.97–13.65 млн лет | |
| Ранний миоцен | Бурдигалский | 20,43–15,97 млн лет |
| Аквитанский | 23.03–20.43 млн лет |
На региональном уровне используются другие системы, основанные на характерных наземных млекопитающих; некоторые из них пересекаются с предшествующим олигоценом и последующими плиоценовыми эпохами:
Возраст европейских наземных млекопитающих
- Туролийский (От 9,0 до 5,3 млн лет)
- Валлезийский (От 11,6 до 9,0 млн лет)
- Астарацкий (От 16,0 до 11,6 млн лет)
- Орлеанский (От 20,0 до 16,0 млн лет)
- Агенский (От 23,8 до 20,0 млн лет)
Возраст наземных млекопитающих в Северной Америке
- Hemphillian (От 10,3 до 4,9 млн лет)
- Кларендонский (От 13,6 до 10,3 млн лет)
- Барстовский (От 16,3 до 13,6 млн лет)
- Хемингфордский (От 20,6 до 16,3 млн лет)
- Арикарийский (От 30,6 до 20,6 млн лет)
Возраст наземных млекопитающих в Южной Америке
- Montehermosan (От 6,8 до 4,0 млн лет)
- Huayquerian (От 9,0 до 6,8 млн лет)
- Майо (От 11,8 до 9,0 млн лет)
- Лавентан (От 13,8 до 11,8 млн лет)
- Colloncuran (От 15,5 до 13,8 млн лет)
- Фриасский (От 16,3 до 15,5 млн лет)
- Сантакрейцерский (От 17,5 до 16,3 млн лет)
- Colhuehuapian (От 21,0 до 17,5 млн лет)
Палеогеография
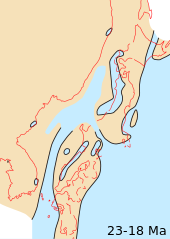
Континенты продолжали дрейф к их нынешним позициям. Из современных геологических особенностей только сухопутный мост между Южная Америка и Северная Америка отсутствовал, хотя Южная Америка приближалась к западному зона субдукции в Тихий океан, вызывая как рост Анды и расширение на юг Мезоамериканский полуостров.
Горное строительство происходило в западной Северная Америка, Европа, и Восточная Азия. Как континентальные, так и морские миоценовые отложения распространены во всем мире, а морские обнажения встречаются у современных береговых линий. Хорошо изученные континентальные обнажения происходят в Северной Америке. Большие равнины И в Аргентина.
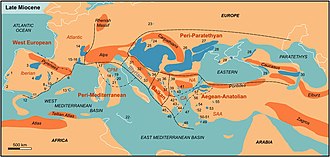
Индия продолжал сталкиваться с Азия, создавая драматические новые Горные хребты. В Тетис морской путь продолжал сокращаться, а затем исчез, когда Африка столкнулся с Евразия в турецкий –Арабский область между 19 и 12 млн лет назад. Последующие поднятие гор на западе Средиземноморье регион и глобальное падение уровня моря в совокупности привели к временному высыханию Средиземного моря (известного как Мессинский кризис солености ) ближе к концу миоцена.
Глобальная тенденция заключалась в увеличении засушливости, вызванной главным образом глобальным похолоданием, снижающим способность атмосферы поглощать влагу. Подъем Восточная Африка в позднем миоцене частично ответственен за сокращение тропические леса в этом регионе, и Австралия стал суше, когда он вошел в зону с малым количеством осадков в позднем миоцене.
Южная Америка
Вовремя Олигоцен и раннего миоцена побережье северной Бразилии,[12] Колумбия, южно-центральное Перу, центральное Чили и большие участки внутренних Патагония подвергались морской проступок.[13] Считается, что нарушения на западном побережье Южной Америки вызваны региональным явлением, в то время как неуклонный рост центральный сегмент Анд представляет собой исключение.[13] Хотя существует множество регистров олиго-миоценовых трансгрессий по всему миру, сомнительно, чтобы они коррелировали.[12]
Считается, что олиго-миоценовая трансгрессия в Патагония мог временно связать Тихий и Атлантический океаны, как следует из находок окаменелостей морских беспозвоночных атлантического и тихоокеанского происхождения в Формация Ла-Каскада.[14][15] Связь произошла бы через узкую эпиконтинентальные морские пути которые сформировали каналы в рассеченная топография.[14][16]
В Антарктическая плита начал подчинять под Южной Америкой 14 миллионов лет назад в миоцене, образуя Чили тройной перекресток. Сначала Антарктическая плита погрузилась только в самой южной оконечности Патагонии, а это означало, что Тройное соединение Чили лежало недалеко от Магелланов пролив. Поскольку южная часть Плита Наска и Chile Rise были поглощены субдукцией, более северные регионы Антарктической плиты начали субдукцию под Патагонией, так что Чилийское тройное соединение со временем продвинулось на север.[17] В астеносферное окно связанных с тройным стыком, нарушил предыдущие модели мантийная конвекция под Патагонией, вызывая подъем ок. 1 км, который полностью изменил трансгрессию олигоцена и миоцена.[16][18]
Как южный Роза Анд в среднем миоцене (14–12 млн лет назад) образовавшиеся тень дождя возникла Патагонская пустыня на восток.[19]
Климат
Климат оставался умеренно теплым, хотя медленное глобальное похолодание, которое в конечном итоге привело к Плейстоцен оледенения продолжение.
Хотя долгосрочная тенденция к похолоданию шла полным ходом, есть свидетельства теплого периода в миоцене, когда глобальный климат соперничал с климатом Олигоцен. Потепление миоцена началось 21 миллион лет назад и продолжалось до 14 миллионов лет назад, когда глобальные температуры резко упали - Переходный климат среднего миоцена (MMCT). К 8 миллионам лет назад температура снова резко упала, и Антарктический ледяной покров уже приближался к нынешним размерам и толщине. Гренландия возможно, начали образовывать большие ледники еще 7-8 миллионов лет назад,[нужна цитата ] хотя климат по большей части оставался достаточно теплым, чтобы поддерживать леса в плиоцене.
Жизнь
Жизнь в миоценовую эпоху в основном поддерживалась двумя новообразованными биомами, лесами ламинарии и лугами. Пастбища позволяют больше пастбищ, таких как лошади, носороги и бегемоты. К концу этой эпохи существовало девяносто пять процентов современных заводов.
Флора

В коэволюция из песчаный, волокнистый, огнестойкий травы и длинноногие общительный копытные с зубы с высокими коронками, привело к значительному расширению травоядных экосистемы, с бродячими стадами больших, быстрый травоядные преследуемый хищники через широкие просторы открытых луга, вытеснив пустыню, лес и браузеры.
Чем выше содержание органики и удержание воды, тем глубже и богаче пастбищные почвы, с долгосрочным захоронение углерода в отложениях образовался углерод и водяной пар раковина. Это в сочетании с более высокой поверхностью альбедо и ниже эвапотранспирация пастбищ, способствовал более прохладному и сухому климату.[21] C4 травы, способные ассимилировать углекислый газ и поливать эффективнее, чем C3 травы, разрослись и стали экологически значимыми ближе к концу миоцена, между 6 и 7 миллионами лет назад.[22] Расширение лугов и излучения среди наземных травоядных коррелирует с колебаниями CO2.[23]
Cycads Между 11,5 и 5 миллионами лет назад, после предыдущего снижения разнообразия из-за климатических изменений, они начали вновь диверсифицироваться, и, таким образом, современные саговники не являются хорошей моделью для «живых ископаемых».[24] Эвкалипт ископаемые листья встречаются в миоцене Новая Зеландия, где род не является сегодня аборигенным, но был завезен из Австралия.[25]
Фауна

И морской, и континентальный фауна были довольно современными, хотя морские млекопитающие были менее многочисленными. Только в изолированных Южной Америке и Австралии существовала сильно различающаяся фауна.
В раннем миоцене несколько олигоценовых групп все еще были разнообразны, в том числе нимравиды, энтелодонты, и трехпалые лошадиные. Как и в предыдущую эпоху олигоцена, ореодонты были все еще разнообразны, но исчезли в самом раннем плиоцене. В течение позднего миоцена млекопитающие были более современными, с легко узнаваемыми псовые, медведи, проциониды, лошадиные, бобры, олень, верблюды, и киты, наряду с ныне вымершими группами, такими как борофагиновые псовые, определенный гомфотеры, трехпалые лошади, а безрогие носороги любят Teleoceras и Афелопс. Острова начали формироваться между Южной и Северной Америкой в позднем миоцене, что позволило наземным ленивцам, таким как Thinobadistes к айленд-хоп в Северную Америку. Расширение богатый кремнеземом C4 травы привели к всемирному исчезновению травоядных видов без зубы с высокими коронками.[26]
Несколько базальных групп млекопитающих дожили до этой эпохи на южных массивах суши, в том числе в Южной Америке. дриолестоид Некролесты и гондана Патагония и Новая Зеландия с Млекопитающее Святого Батана. Немногочисленные метатерии все еще были, например, американские и евразийские герпеттерииды и перадектиды Такие как Siamoperadectes, и южноамериканский Спарассодонты.
Однозначно узнаваемый веселящиеся утки, ржанки, типичные совы, какаду и вороны появляются в миоцене. К концу эпохи все или почти все современные группы птиц, как полагают, присутствовали; несколько окаменелостей птиц после миоцена, которые нельзя с полной уверенностью поместить в эволюционное древо, просто слишком плохо сохранились, а не слишком двусмысленны по своему характеру. В эту эпоху морские птицы достигли своего наивысшего разнообразия.
Примерно 100 видов обезьяны жили в это время по всей Африке, Азии и Европе и сильно различались по размеру, диете и анатомии. Из-за скудных окаменелостей неясно, какие обезьяны способствовали развитию современного гоминид clade, но молекулярные данные указывают на то, что эта обезьяна жила от 7 до 8 миллионов лет назад.[27] Первый гоминины (двуногий человекообразные обезьяны) появились в Африке в самом конце миоцена, в том числе Сахелантроп, Оррорин, и ранняя форма Ардипитека (А. кадабба ) расхождение между шимпанзе и человеком считается, что это произошло в это время.[28]
Расширение пастбищ в Северной Америке также привело к взрывной радиации среди змей.[29] Раньше змеи были второстепенным компонентом фауны Северной Америки, но в миоцене количество видов и их распространенность резко возросли с появлением первых животных. гадюки и elapids в Северной Америке и значительная диверсификация Colubridae (включая происхождение многих современных родов, таких как Неродиа, Лампропелтис, Питуофис и Пантерофис ).[29]


В океанах бурые водоросли, называется ламинария, размножается, поддерживая новые виды морской жизни, в том числе выдры, рыбы и различные беспозвоночные.
Китообразные достигли наибольшего разнообразия в миоцене,[30] с более чем 20 признанными родами усатые киты по сравнению только с шестью живыми родами.[31] Эта диверсификация коррелирует с появлением гигантских макрохищников, таких как мегазубые акулы и хищные птицы. кашалоты.[32] Яркими примерами являются C. мегалодон и L. melvillei.[32] Другие известные крупные акулы были С. chubutensis, Isurus hastalis, и Hemipristis serra.
Крокодилы также показали признаки диверсификации в миоцене. Самой крупной формой среди них был гигантский кайман Пуруссавр который населял Южную Америку.[33] Другой гигантской формой был ложный гавиал Рамфосух, который населял современную эпоху Индия. Странная форма, Mourasuchus также процветал рядом с Пуруссавр. Этот вид разработал специализированный механизм фильтрационного питания и, вероятно, охотился на небольшую фауну, несмотря на свои гигантские размеры.
В ластоногие, появившийся ближе к концу олигоцена, стал более водным. Видный род был Аллодесмус.[34] Свирепый морж, Пелагиарктос могли охотиться на другие виды ластоногих, включая Аллодесмус.
Более того, южноамериканец воды засвидетельствовали прибытие Megapiranha paranensis, которые были значительно больше современности пираньи.
Новая Зеландия Летопись окаменелостей миоцена России особенно богата. Морские отложения демонстрируют множество китообразные и пингвины, иллюстрирующий эволюцию обеих групп в современных представителей. Ранний миоцен Фауна Сент-Батанса является единственной кайнозойской земной летописью окаменелостей суши, демонстрирующей большое разнообразие не только птица видов, в том числе ранних представителей клад, таких как моа, киви и адзевиллы, но и разнообразная герпетофауна sphenodontians, крокодилы и черепаха а также богатая фауна наземных млекопитающих, состоящая из различных видов летучие мыши и загадочный Млекопитающее святого Батана.
Океаны
Имеются данные изотопов кислорода на Программа глубоководного бурения участки, где лед начал образовываться в Антарктиде около 36 млн лет назад во время эоцен. Дальнейшее заметное снижение температуры во время Средний миоцен на 15 млн лет назад, вероятно, отражает усиление роста льда в Антарктиде. Таким образом, можно предположить, что в Восточной Антарктиде было несколько ледников в период от раннего до среднего миоцена (23–15 млн лет назад). Океаны охлаждались частично из-за образования Антарктическое циркумполярное течение, а около 15 миллионов лет назад ледяная шапка в южном полушарии начала увеличиваться до нынешней формы. Ледяная шапка Гренландии образовалась позже, в Средний плиоцен время, около 3 миллионов лет назад.
Нарушение среднего миоцена
«Разрушение среднего миоцена» относится к волне вымирания наземных и водных форм жизни, которые возникли после миоценового климатического оптимума (18-16 млн лет назад), примерно 14,8-14,5 миллионов лет назад, во время Langhian этап среднего миоцена. Между 14,8 и 14,1 млн лет назад произошел крупный и постоянный этап похолодания, связанный с увеличением образования холодных антарктических глубинных вод и значительным ростом ледникового покрова Восточной Антарктики. Средний миоцен δ18Увеличение O, то есть относительное увеличение более тяжелого изотопа кислорода, было отмечено в Тихом океане, Южном океане и Южной Атлантике.[35]
Событие удара
Большой ударное событие произошли либо в миоцене (23–5,3 млн лет), либо в плиоцене (5,3–2,6 млн лет). Событие сформировало Кратер Каракуль (Диаметр 52 км), возраст которого оценивается менее 23 млн лет.[36] или менее 5 млн лет.[37]
Смотрите также
- Шкала геологического времени
- Список ископаемых
- Категория: Животные миоцена
Рекомендации
- ^ Krijgsman, W .; Garcés, M .; Langereis, C.G .; Daams, R .; Van Dam, J .; Van Der Meulen, A.J .; Agustí, J .; Кабрера, Л. (1996). «Новая хронология континентальных летописей среднего и позднего миоцена в Испании». Письма по науке о Земле и планетах. 142 (3–4): 367–380. Bibcode:1996E и PSL.142..367K. Дои:10.1016 / 0012-821X (96) 00109-4.
- ^ Реталлак, Дж. Дж. (1997). «Неогеновая экспансия Североамериканских прерий». ПАЛАИ. 12 (4): 380–390. Дои:10.2307/3515337. JSTOR 3515337. Получено 2008-02-11.
- ^ Ван Куверинг, Джон; Кастрадори, Давиде; Сита, Мария; Хильген, Фредерик; Рио, Доменико (сентябрь 2000 г.). «Основание занклинского яруса и плиоценовой серии» (PDF). Эпизоды. 23: 179–187.
- ^ «Миоцен». Dictionary.com Несокращенный. Случайный дом.
- ^ «Миоцен». Словарь Merriam-Webster.
- ^ Видеть:
- Письмо от Уильям Уэвелл к Чарльз Лайель от 31 января 1831 г. в: Тодхантер, Исаак, изд. (1876 г.). Уильям Уэвелл, доктор медицины, магистр Тринити-колледжа, Кембридж: отчет о его трудах с отрывками из его литературной и научной переписки. т. 2. Лондон, Англия: Macmillan and Co. p. 111.
- Лайель, Чарльз (1833). Принципы геологии,…. т. 3. Лондон, Англия: Джон Мюррей. п. 54. С п. 54: «Следующую предшествующую третичную эпоху мы назовем миоценом, от μειων, minor и χαινος, Recens, лишь небольшая часть ископаемых раковин, заключенных в формациях этого периода, которые относятся к недавним видам».
- ^ «Миоцен». Интернет-словарь этимологии. Получено 2016-01-20.
- ^ Лайель, Чарльз (1833). Принципы геологии,…. т. 3. Лондон, Англия: Джон Мюррей. п. 54.
- ^ "BBC Nature - видео, новости и факты эпохи миоцена". BBC. Получено 2016-11-13.
- ^ Чжишэн, Ань; Куцбах, Джон Э .; Prell, Warren L .; Портер, Стивен К. (3 мая 2001 г.). «Эволюция азиатских муссонов и поэтапное поднятие Гималайско-Тибетского плато со времен позднего миоцена». Природа. 411 (6833): 62–66. Bibcode:2001Натура.411 ... 62Z. Дои:10.1038/35075035. PMID 11333976. S2CID 4398615.
- ^ Роберт А. Роде (2005). «База данных GeoWhen». Получено 8 марта, 2011.
- ^ а б Россетти, Дилче Ф .; Безерра, Франсиско Х.Р .; Домингес, Хосе М.Л. (2013). «Позднеолигоцен-миоценовые трансгрессии вдоль экваториальной и восточной окраин Бразилии». Обзоры наук о Земле. 123: 87–112. Bibcode:2013ESRv..123 ... 87R. Дои:10.1016 / j.earscirev.2013.04.005.
- ^ а б Мачаре, Хосе; Деврис, Томас; Бэррон, Джон; Фуртанье, Элизабет (1988). «Олиго-миоценовая трансгрессия вдоль окраины Пацифи в Южной Америке: новые палеонтологические и геологические свидетельства из бассейна Писко (Перу)» (PDF). Геодинамика. 3 (1–2): 25–37.
- ^ а б Энсинас, Альфонсо; Перес, Фелипе; Нильсен, Свен; Finger, Kenneth L .; Валенсия, Виктор; Duhart, Пол (2014). «Геохронологические и палеонтологические свидетельства связи между Тихим океаном и Атлантикой в течение позднего олигоцена – раннего миоцена в Патагонских Андах (43–44 ° ю.ш.)». Журнал южноамериканских наук о Земле. 55: 1–18. Bibcode:2014JSAES..55 .... 1E. Дои:10.1016 / j.jsames.2014.06.008. HDL:10533/130517.
- ^ Нильсен, С. (2005). «Кайнозойские Strombidae, Aporrhaidae и Struthiolariidae (Gastropoda, Stromboidea) из Чили: их значение для биогеографии фауны и климата юго-восточной части Тихого океана». Журнал палеонтологии. 79: 1120–1130. Дои:10.1666 / 0022-3360 (2005) 079 [1120: csaasg] 2.0.co; 2.
- ^ а б Гийам, Бенджамин; Мартинод, Джозеф; Хассон, Лоран; Роддаз, Мартин; Рикельме, Родриго (2009). «Неогеновое поднятие центрально-восточной Патагонии: динамический ответ на активное субдукцию спредингового хребта?». Тектоника. 28.
- ^ Cande, S.C .; Лесли, Р. Б. (1986). «Позднекайнозойская тектоника Южно-Чилийского желоба». Журнал геофизических исследований: твердая Земля. 91 (B1): 471–496. Bibcode:1986JGR .... 91..471C. Дои:10.1029 / jb091ib01p00471.
- ^ Гийом, Бенджамин; Гаутерон, Сесиль; Симон-Лабрик, Тибо; Мартинод, Джозеф; Роддаз, Мартин; Дувиль, Эрик (2013). «Управление динамической топографией на эволюции патагонского рельефа на основе низкотемпературной термохронологии». Письма по науке о Земле и планетах. 3: 157–167. Bibcode:2013E и PSL.364..157G. Дои:10.1016 / j.epsl.2012.12.036.
- ^ Фольгера, Андрес; Энсинас, Альфонсо; Эчауррен, Андрес; Джанни, Гвидо; Ортс, Дарио; Валенсия, Виктор; Карраско, Габриэль (2018). «Ограничения на неогеновый рост центральных Патагонских Анд на широте тройного сочленения Чили (45–47 ° ю.ш.) с использованием геохронологии U / Pb в синорогенных пластах». Тектонофизика. 744: 134–154. Дои:10.1016 / j.tecto.2018.06.011. HDL:11336/88399.
- ^ Attorre, F .; Francesconi, F .; Талеб, Н .; Scholte, P .; Saed, A .; Alfo, M .; Бруно, Ф. (2007). «Переживет ли драконья кровь в следующий период изменения климата? Текущее и будущее потенциальное распространение Dracaena cinnabari (Сокотра, Йемен)». Биологическое сохранение. 138 (3–4): 430–439. Дои:10.1016 / j.biocon.2007.05.009.
- ^ Retallack, Грегори (2001). «Кайнозойское расширение пастбищ и похолодание климата» (PDF). Журнал геологии. Издательство Чикагского университета. 109 (4): 407–426. Bibcode:2001JG .... 109..407R. Дои:10.1086/320791. S2CID 15560105. Архивировано из оригинал (PDF) на 2013-05-06.
- ^ Osborne, C.P .; Бирлинг, Д.Дж. (2006). «Зеленая революция природы: выдающийся эволюционный рост растений C4». Философские труды Королевского общества B: биологические науки. 361 (1465): 173–194. Дои:10.1098 / rstb.2005.1737. ЧВК 1626541. PMID 16553316.
- ^ Вольфрам М. Кюршнер, Златко Квачек и Дэвид Л. Дилчер (2008). «Влияние миоценовых колебаний содержания углекислого газа в атмосфере на климат и эволюцию наземных экосистем». Труды Национальной академии наук. 105 (2): 449–53. Bibcode:2008ПНАС..105..449К. Дои:10.1073 / pnas.0708588105. ЧВК 2206556. PMID 18174330.
- ^ Сюзанна С. Реннер (2011). «Живое ископаемое моложе мысли». Наука. 334 (6057): 766–767. Bibcode:2011Наука ... 334..766R. Дои:10.1126 / science.1214649. PMID 22076366. S2CID 206537832.
- ^ «Окаменелости эвкалипта в Новой Зеландии - тонкий конец клина - Майк Поул». 2014-09-22.
- ^ Стивен М. Стэнли (1999). История системы Земля. Нью-Йорк: Фриман. С. 525–526. ISBN 0-7167-2882-6.
- ^ Йирка, Боб (15 августа 2012 г.). «Новые генетические данные показывают, что люди и человекообразные обезьяны разошлись раньше, чем предполагалось». Phys.org.
- ^ Бегун, Дэвид. «Окаменелости миоценовых гоминоидов» (PDF). Университет Торонто. Архивировано из оригинал (PDF) 30 октября 2013 г.. Получено 11 июля, 2014.
- ^ а б Холман, Дж. Алан (2000). Ископаемые змеи Северной Америки (Первое изд.). Блумингтон, Индиана: Издательство Индианского университета. С. 284–323. ISBN 0253337216.
- ^ Питер Климли и Дэвид Эйнли (1996). Большие белые акулы: биология Carcharodon carcharias. Академическая пресса. ISBN 0-12-415031-4. Архивировано из оригинал на 2012-10-12. Получено 2011-08-12.
- ^ Dooley, Alton C .; Fraser, Nicholas C .; Ло, Чжэ-Си (2004). «Самый ранний известный представитель клады рорквалов - серых китов (Mammalia, Cetacea)». Журнал палеонтологии позвоночных. 24 (2): 453–463. Дои:10.1671/2401. ISSN 0272-4634. S2CID 84970052.
- ^ а б Оливье Ламбер; Джованни Биануччи; Клаас Пост; Кристиан де Мюисон; Родольфо Салас-Гисмонди; Марио Урбина; Джелле Реумер (2010). «Гигантский укус нового хищного кашалота миоценовой эпохи Перу». Природа. 466 (7302): 105–108. Bibcode:2010Натура.466..105л. Дои:10.1038 / природа09067. PMID 20596020. S2CID 4369352.
- ^ Оранджел А. Агилера, Дуглас Рифф и Жан Боккантен-Вильянуэва (2006). "Новый гигант Пусуссавр (Crocodyliformes, Alligatoridae) из формации Urumaco верхнего миоцена, Венесуэла " (PDF). Журнал систематической палеонтологии. 4 (3): 221–232. Дои:10.1017 / S147720190600188X. S2CID 85950121. Архивировано из оригинал (PDF) на 2012-03-29.
- ^ Лоуренс Дж. Барнс и Киёхару Хирота (1994). «Миоценовые ластоногие отарид подсемейства Allodesminae в северной части Тихого океана: систематика и взаимосвязи». Островная арка. 3 (4): 329–360. Дои:10.1111 / j.1440-1738.1994.tb00119.x.
- ^ Кеннет Г. Миллер и Ричард Г. Фэрбенкс (1983). «Свидетельства изменений абиссальной циркуляции олигоцена-среднего миоцена в западной части Северной Атлантики». Природа. 306 (5940): 250–253. Bibcode:1983Натура.306..250М. Дои:10.1038 / 306250a0. S2CID 4337071.
- ^ Bouley, S .; Baratoux, D .; Baratoux, L .; Colas, F .; Dauvergne, J .; Лосяк, А .; Vaubaillon, J .; Bourdeille, C .; Jullien, A .; Ибадинов, К. (Американский геофизический союз, осеннее собрание 2011 г.) (2011). «Каракуль: молодой сложный ударный кратер на Памире, Таджикистан». Тезисы осеннего собрания AGU. 2011: P31A – 1701. Bibcode:2011AGUFM.P31A1701B.CS1 maint: использует параметр авторов (связь)
- ^ Гуров Е.П., Гурова Е.П., Ракицкая Р.Б., Ямниченко А.Ю. (1993) (1993). «Каракульская впадина на Памире - первое ударное сооружение в Центральной Азии» (PDF). Луна и планетология XXIV, стр. 591-592: 591. Bibcode:1993ЛПИ .... 24..591Г.CS1 maint: использует параметр авторов (связь)
дальнейшее чтение
- Кокс, К. Барри и Мур, Питер Д. (1993): Биогеография. Экологический и эволюционный подход (5-е изд.). Научные публикации Блэквелла, Кембридж. ISBN 0-632-02967-6
- Огг, Джим (2004): "Обзор разрезов и точек стратотипа глобальной границы (GSSP) ". Проверено 30 апреля 2006.