Homo heidelbergensis - Homo heidelbergensis
| Homo heidelbergensis | |
|---|---|
 | |
| В типовой образец Мауэр 1 | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Haplorhini |
| Инфразаказ: | Симиобразные |
| Семья: | Гоминиды |
| Подсемейство: | Гоминины |
| Племя: | Гоминини |
| Род: | Гомо |
| Разновидность: | †Х. heidelbergensis |
| Биномиальное имя | |
| †Homo heidelbergensis Schoetensack, 1908 | |
| Синонимы | |
| |
Homo heidelbergensis (также H. sapiens heidelbergensis) является вымершим разновидность или же подвид из архаичный человек которые существовали в Средний плейстоцен. Он был включен в подвид H. erectus в 1950 году как Он. Heidelbergensis, но ближе к концу века он стал более широко классифицироваться как отдельный вид. Обсуждается вопрос о том, следует ли ограничивать этот вид только Европой или включать также африканские и азиатские образцы, и это еще больше осложняется тем, что типовой образец (Мауэр 1 ) является челюстной костью, потому что челюстные кости имеют мало диагностических признаков и обычно отсутствуют среди образцов среднего плейстоцена. Таким образом, обсуждается, могут ли некоторые из этих образцов быть разделены на их собственные виды или подвиды H. erectus. Поскольку классификация так спорна, средний плейстоцен часто называют «путаницей посередине».
Х. heidelbergensis рассматривается как хроновиды, произошедший от африканской формы H. erectus (иногда называют Х. эргастер ), и по соглашению помещается как самый последний общий предок между современными людьми (Х. сапиенс или же Х. с. sapiens) и Неандертальцы (H. neanderthalensis или же Х. с. неандертальцы). Многие образцы отнесены к Х. heidelbergensis вероятно, существовали и после раскола современных людей и неандертальцев. В среднем плейстоцене размер мозга в среднем составлял около 1200 куб. См, что сопоставимо с размером мозга современного человека. Высоту в среднем плейстоцене можно оценить только по останкам трех местонахождений: Sima de los Huesos, Испания - 169,5 см (5 футов 7 дюймов) для мужчин и 157,7 см (5 футов 2 дюйма) для женщин; 165 см (5 футов 5 дюймов) для женщин от Джиннюшань, Китай; и 181,2 см (5 футов 11 дюймов) для образца из Кабве, Замбия. В целом они кажутся довольно высокими. Как и у неандертальцев, у них была широкая грудь и крепкий.
В среднем плейстоцене Африки и Европы наступает период позднего Ашельский технологии, расходящиеся с более ранними и современными H. erectus, и, вероятно, связано с повышением интеллекта. Огонь, вероятно, стал неотъемлемой частью повседневной жизни после 400000 лет назад, и это примерно совпадает с более постоянным и широко распространенным заселением Европы (см. Выше 45 ° с.ш. ), а появление рывок технология создания копий. Возможно, они были в состоянии осуществлять скоординированные стратегии охоты, и, похоже, у них была более высокая зависимость от мяса.
Таксономия
История исследований
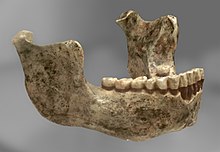
Первая окаменелость, Мауэр 1 (челюстная кость), был обнаружен рабочим в Мауэр, к юго-востоку от Гейдельберг, Германия, в 1907 году. В следующем году он был официально описан немецким антропологом. Отто Шотенсак, кто сделал это типовой образец нового вида, Homo heidelbergensis.[1] Он создал новый вид в первую очередь из-за архаичности нижней челюсти - в частности, ее огромного размера - и это была самая старая человеческая челюсть в европейской летописи окаменелостей, возраст которой составлял 640 000 лет. Нижняя челюсть хорошо сохранилась, отсутствуют только левые премоляры, часть 1-го левого моляра, кончик левого моляра. венечный отросток (на шарнире челюсти), а также фрагменты средней части челюсти, так как челюсть была обнаружена в 2 частях и их нужно было склеить. Возможно, он принадлежал молодому человеку из-за легкого износа 3-го моляра.[2]
Мауэр 1 был связан с Кромериан промышленность. В 1921 году череп Кабве 1 был обнаружен швейцарским шахтером Томом Цвигларом в Кабве, Замбия (в то время Брокен Хилл, Северная Родезия ), и был отнесен к новому виду ",H. rhodesiensis", английский палеонтолог Артур Смит Вудворд.[3] Это был один из многих предполагаемых видов Средний плейстоцен Гомо которые были описаны на протяжении первой половины 20 века. В 1950 г. Эрнст Майр вошел в сферу антропологии и, изучив «ошеломляющее разнообразие названий», решил определить только 3 вида Гомо: "H. transvaalensis"( австралопитеки ), H. erectus (включая нижнюю челюсть Мауэра и различные предполагаемые африканские и азиатские таксоны), и Homo sapiens (включая все, что младше H. erectus, таких как современные люди и Неандертальцы ). Майр определил их как последовательную линию, в которой каждый вид эволюционировал в следующий (хроновиды ). Хотя позже Майр изменил свое мнение об австралопитеках (признав Австралопитек ), его более консервативный взгляд на архаичный человек разнообразие получило широкое распространение в последующие десятилетия.[4]

Хотя H. erectus до сих пор сохраняется как сильно изменчивый, широко распространенный и долговечный вид, все еще много споров о том, оправдано ли погружение в него всех остатков среднего плейстоцена. Комкование Майра Х. heidelbergensis впервые выступил против американского антрополога Фрэнсис Кларк Хауэлл в 1960 г. В 1974 г. британский физический антрополог Крис Стрингер указал на сходство между Кабве 1 и греческим Петралона черепа к черепам современного человека (Х. сапиенс или же Х. с. sapiens) и неандертальцы (H. neanderthalensis или же Х. с. неандертальцы). Итак, Стрингер назначил их Homo sapiens Sensu lato («в широком смысле»), как предок современных людей и неандертальцев. В 1979 году Стрингер и финский антрополог Бьорн Куртен обнаружили, что черепа Кабве и Петралона связаны с кромерианами, как нижняя челюсть Мауэра, и, таким образом, предположили, что эти 3 популяции могут быть связаны друг с другом. Хотя эти окаменелости плохо сохранились и не обеспечивают многих сопоставимых возможных диагностических признаков (и аналогично в то время было трудно правильно определить уникальный вид), они утверждали, что, по крайней мере, эти образцы среднего плейстоцена должны быть отнесены к Х. (s.?) Heidelbergensis или же "H. (s.?) Rhodesiensis«(в зависимости, соответственно, от включения или исключения нижней челюсти Мауэра) формально признать их сходство.[5]
Дальнейшая работа наиболее влиятельна Стрингера, палеоантрополога. Ян Таттерсолл, а биолог-эволюционист Филлип Райтмайр сообщил о дальнейших различиях между афро-европейскими образцами среднего плейстоцена и H. erectus Sensu stricto («в строгом смысле», в данном случае образцы из Восточной Азии).[6] Следовательно, афро-европейские останки от 600 до 300 тысяч лет назад - в первую очередь от Кабве, Петралоны, Бодо, и Араго - часто классифицируются как Х. heidelbergensis. В 2010 году американский физический антрополог Джеффри Х. Шварц и Таттерсолл предложил классифицировать все европейские и азиатские образцы среднего плейстоцена, а именно из Дали и Джиннюшань в Китае - как Х. heidelbergensis.[4] Эта модель не так общепринята. После идентификации в 2010 году генетического кода некоторых уникальных архаичных человеческих видов в Сибири, названных "Денисовцы "ожидающие диагностических находок окаменелостей, предполагается, что азиатские останки могут представлять тот же самый вид.[5] Таким образом, азиатские образцы среднего плейстоцена, такие как человек Дали или индийский Нармада Ман, оставайтесь загадочными.[7] Институт палеонтологии при г. Гейдельбергский университет, где нижняя челюсть Мауэра хранилась с 1908 г., изменила этикетку с Он. Heidelbergensis к Х. heidelbergensis в 2015 году.[8]

В 1976 г. Sima de los Huesos (SH) в Сьерра-де-Атапуэрка, Испания, испанские палеонтологи Эмилиано Агирре, Хосе Мария Басабе и Тринидад Торрес начали раскопки архаических человеческих останков. Их исследование сайта было вызвано обнаружением нескольких нести останки (Ursus deningeri ) с начала 20 века спелеологами-любителями (которые впоследствии уничтожили некоторые человеческие останки в этом разделе). К 1990 году было зарегистрировано около 600 человеческих останков, а к 2004 году их количество увеличилось примерно до 4000. Это не менее 28 человек, из которых, возможно, только 1 ребенок, а остальные - подростки и молодые люди. Сборка окаменелостей исключительно полная, трупы целиком захоронены быстро, со всеми элементами тела.[9] В 1997 году испанский палеоантрополог Хуан Луис Арсуага назначил их Х. heidelbergensis, но в 2014 году он отказался от этого, заявив, что неандертальские черты, присутствующие в нижней челюсти Мейра, отсутствуют у людей SH.[10]
Классификация
В палеоантропологии Средний плейстоцен часто называют "путаницей посередине", потому что классификация видов на уровне архаичный человек Остатки этого периода времени активно обсуждались. Предки современного человека (Homo sapiens или же Х. с. sapiens) и неандертальцы (H. neanderthalensis или же Х. с. неандертальцы) разошлись в этот период времени, и, по соглашению, Х. heidelbergensis обычно считается LCA. Это сделало бы Х. heidelbergensis член хроновиды.[11][7] Это много споров, если имя Х. heidelbergensis можно распространить на людей среднего плейстоцена по всему Старому Свету, или, если лучше, ограничить это только Европой. В последнем случае африканские останки среднего плейстоцена можно разделить на "H. rhodesiensis"или" архаичный Х. сапиенс".[12][13][14][15] Согласно последнему мнению "H. rhodesiensis"может рассматриваться как прямой предок современного человека, или как"H. helmei "которые превратились в современных людей.[16]
Что касается европейских останков среднего плейстоцена, то некоторые из них более прочно относятся к линии неандертальцев (а именно SH, Pontnewyyd, Steinheim, и Swanscombe ), в то время как другие, кажется, имеют несколько уникальных неандертальских черт (Араго, Чепрано, Vértesszőlős, Bilzingsleben, Мала Баланица, и Ароейра ). Из-за этого предполагается, что в этом регионе и в этот период времени существовало несколько линий (или видов), но французский палеоантрополог Жан-Жак Хюблен считает это неоправданной экстраполяцией, поскольку они могли быть просто разными, но все же взаимосвязанными популяциями одного, очень изменчивого вида.[17] Менее производный материал также может быть разделен на их собственные виды или подвиды H. erectus s. л. (например, материал Arago как "Он. Tautavelensis").[6] Нет четкого различия между последними потенциальными Х. heidelbergensis материал - в частности, Steinheim и SH - и самые ранние неандертальские образцы -Biache, Франция; Ehringsdorf, Германия; или же Saccopastore, Италия.[18]
Использование нижней челюсти Мауэра, изолированной челюстной кости, в качестве типовой образец для этого вида было проблематично, так как нижняя челюсть отсутствует у нескольких образцов среднего плейстоцена, и в любом случае она не имеет многих диагностических признаков. Антрополог Уильям Штраус сказал по этой теме: «В то время как череп - творение Бога, челюсть - это работа дьявола». Если нижняя челюсть Мауэра на самом деле принадлежит к другому виду, нежели череп Кабве и большинство других афро-европейских архаичных людей среднего плейстоцена, то "H. rhodesiensis" взял бы приоритет как название LCA.[7]
Эволюция
Что касается его эволюции, Х. heidelbergensis считается, что произошел от африканского H. erectus- иногда классифицируется как Homo ergaster —В течение первого ранние экспансии гомининов из Африки началось примерно 2 миллиона лет назад. Те, кто расселились по Европе и остались в Африке, превратились в Х. heidelbergensis или указаны в Х. heidelbergensis в Европе и "H. rhodesiensis"в Африке, а те, что рассредоточились по Восточной Азии, превратились в H. erectus s. с.[2] Точное происхождение от вида-предка неясно из-за большого пробела в летописи окаменелостей человека ближе к концу Ранний плейстоцен.[16] В 2016 году Антонио Профико и его коллеги предположили, что черепные материалы возрастом 875000 лет с участка Гомборе II Мелка Кунтуре Формация, Эфиопия, представляет собой переходный морф между Х. эргастер и Х. heidelbergensis, и таким образом постулировал, что Х. heidelbergensis возник в Африке, а не в Европе.[16]

Согласно генетическому анализу, последний общий предок (LCA) современных людей и неандертальцев разделились на современную человеческую линию и линию неандертальцев / денисовцев, а последние позже разделились на неандертальцев и денисовцев. В соответствии с ядерная ДНК Анализ показывает, что люди SH, возрастом 430000 лет, более тесно связаны с неандертальцами, чем с денисовцами, что позволяет предположить, что современный человек / неандерталец (LCA) существовал задолго до того, как многие европейские образцы, обычно относящиеся к Х. heidelbergensis сделал, например, материалы Араго и Петралона.[19]
В 1997 году испанский археолог Хосе Мария Бермудес де Кастро, Арсуага и его коллеги описали возраст примерно миллион лет H. предшественник из Гран Долина, Сьерра-де-Атапуэрка, и предложил вытеснить этот вид вместо Х. heidelbergensis для LCA между современными людьми и неандертальцами, с Х. heidelbergensis происходящий от него и являющийся строго европейским видом, предком только неандертальцам.[20] Это было опровергнуто в 2020 году Фридо Велкер и его коллегами, которые проанализировали древние белки, собранные из H. предшественник зуб и обнаружил, что он был членом сестринской линии LCA, а не самим LCA (то есть Х. heidelbergensis не происходило из H. предшественник).[21]
Расселение людей за пределы 45 ° с.ш. кажется, было довольно ограниченным во время Нижний палеолит, с доказательствами кратковременного расселения на север, начавшегося миллион лет назад. Начиная с 700000 лет назад, более постоянные популяции, по-видимому, сохранялись за границей, совпадающей с распространением ручной топор технологии по всей Европе, возможно, связанные с рассредоточением Х. heidelbergensis и изменения в поведении, чтобы справиться с холодным климатом. Такое заселение становится намного более частым после 500 000 лет назад.[22]
Анатомия
Череп

По сравнению с ранним плейстоценом H. erectus/эргастерЛюди среднего плейстоцена имеют гораздо более человеческое лицо. Носовое отверстие расположено в черепе полностью вертикально, а передний носовой порог может быть гребнем или иногда выступающим позвоночником. В режущие каналы (на небо ) открываются около зубов и ориентированы, как у более поздних человеческих видов. В лобная кость широкий, теменная кость можно расширить, а плоскоклеточная часть височной кости высокий и изогнутый, что может быть связано с увеличением размера мозга. В клиновидной кости имеет позвоночник, идущий вниз, а суставной бугорок на нижней стороне черепа может заметно выступать, поскольку поверхность за шарниром челюсти в остальном довольно плоская.[23]
В 2004 году Райтмайр оценил объемы мозга 10 людей среднего плейстоцена, которые по-разному связаны с Х. heidelbergensis- от Кабве, Бодо, Ндуту, Дали, Джинниушан, Петралона, Штайнхейм, Араго и 2 из SH. Этот набор дает средний объем около 1206 см3, в диапазоне от 1100 до 1390 см3. Он также усреднил объем мозга 30 H. erectus/эргастер Образцы, охватывающие почти 1,5 миллиона лет из Восточной Азии и Африки, как 973 куб. см, и, таким образом, завершили значительный скачок в размере мозга, хотя признается, что размер мозга был чрезвычайно изменчивым от 727 до 1231 куб. см в зависимости от периода времени, географического региона, и даже между особями в пределах одной популяции (последнее, вероятно, из-за заметного полового диморфизма, когда самцы намного крупнее самок).[23] Для сравнения, у современных людей размер мозга в среднем составляет 1270 куб. См (78 куб. Дюймов) у мужчин и 1130 куб. Дюймов (69 куб. Дюймов) у женщин;[24] а для неандертальцев 1600 см3 (98 дюймов3) для мужчин и 1300 см3 (79 дюймов3) для женщин.[25][26][27]

В 2009 году палеонтологи Орелиен Мунье, Франсуа Маршаль и Сильвана Кондеми опубликовали первый дифференциальный диагноз Х. heidelbergensis с использованием нижней челюсти Мауэра, а также материала из Тигеннифа, Алжир; SH, Испания; Араго, Франция; и Montmaurin, Франция. Они перечислили такие диагностические признаки как: уменьшенный подбородок, выемка в субментальное пространство (около глотки), параллельные верхняя и нижняя границы нижней челюсти при виде сбоку, несколько подбородочное отверстие (дырочки для сосудов) возле щечных зубов, горизонтальный ретромолярное пространство (щель за коренными зубами), желоб между коренными зубами и ramus (который выступает, чтобы соединиться с черепом), общая длинная челюсть, глубокая ямка (депрессия) для жевательная мышца (закрывающий челюсть) небольшой гониальный угол (угол между телом нижней челюсти и ветвью), обширная плоская альвеолярная (расстояние от передней лунки зуба до задней части челюсти), развитая плоская треугольная поверхность (около шарнира челюсти) и подъязычная линия берущие начало на уровне 3-го моляра.[2]
Строить
Тенденции изменения размеров тела в среднем плейстоцене не видны из-за общего отсутствия костей конечностей и нечерепных (посткраниальных) остатков. Основываясь на длине различных длинные кости, люди SH в среднем составляли примерно 169,5 см (5 футов 7 дюймов) для мужчин и 157,7 см (5 футов 2 дюйма) для женщин, с максимальными размерами соответственно 177 см (5 футов 10 дюймов) и 160 см (5 футов 3 дюйма). Высота частичного скелета самки из Цзиннюшань при жизни оценивается примерно в 165 см (5 футов 5 дюймов), что намного выше, чем у самок SH. Большая большеберцовая кость из Кабве, как правило, составляла 181,2 см (5 футов 11 дюймов) среди самых высоких экземпляров среднего плейстоцена, но возможно, что этот человек был либо необычно большим, либо имел гораздо более длинную большеберцовую кость. бедренная кость соотношение, чем ожидалось. Если эти образцы являются представителями своих континентов, они предполагают, что люди выше среднего и высокого роста преобладали в Среднем плейстоцене Старого Света. Если это так, то большинство популяций любого архаичного человеческого вида в среднем имели бы в среднем 165–170 см (5 футов 5 дюймов – 5 футов 7 дюймов) в высоту. Ранние современные люди были заметно выше, а Схул и Кафзех остаются в среднем 185,1 см (6 футов 1 дюйм) для мужчин и 169,8 см (5 футов 7 дюймов) для женщин, в среднем 177,5 см (5 футов 10 дюймов), возможно, для повышения энергоэффективности дальних путешествий с более длинными ногами .[28]
Человек бауплан (план тела) развился в Х. эргастер, и характеризует все позже Гомо видов, но среди более производных членов есть 2 различных морфа: узкое и изящное телосложение, как у современных людей, и более широкое и крепкое телосложение, как у неандертальцев. Когда-то считалось, что строение неандертальцев было уникальным для неандертальцев на основе изящного Х. эргастер частичный каркас KNM WT-15000 ("Мальчик Туркана "), но открытие некоторых элементов скелета среднего плейстоцена (хотя в целом фрагментарно и далеко и редко), кажется, предполагает, что люди среднего плейстоцена в целом имели более неандертальскую морфу. Таким образом, современный человеческий морф может быть уникальным для современных людей, эволюционируя совсем недавно. Это наиболее ярко демонстрируется в исключительно хорошо сохранившемся комплексе SH. На основании прочности черепа было принято предположение, что люди среднего плейстоцена имели высокую степень половой диморфизм, но люди SH демонстрируют современный человекоподобный уровень.[29]
Люди SH и другие представители среднего плейстоцена Гомо имеют более базальный таз и бедро (больше похоже на предыдущие Гомо чем неандертальцы). В целом широкий и эллиптический таз шире, выше и толще (расширен в переднезаднем направлении), чем у неандертальцев или современных людей, и сохраняет расположенный спереди ацетабулокристаллический упор (который поддерживает гребни подвздошных костей при отведении бедра), хорошо выраженная надбровная борозда (между тазобедренным суставом и подвздошной костью), тонкая и прямоугольная верхняя ветвь лобка (в отличие от толстого и толстого у современных людей). У всех архаичных людей ступня выше блок лодыжки, делая голеностопный сустав более гибким (особенно тыльное и подошвенное сгибание).[29]
Патология
На левой стороне его лица череп SH (Череп 5) представляет собой старейший известный случай орбитальный целлюлит (глазная инфекция, развившаяся из абсцесс во рту). Вероятно, это вызвало сепсис, убивая человека.[30][31][32]
Мужской таз SH (таз 1), основанный на дегенерации сустава, может прожить более 45 лет, что делает его одним из старейших примеров этой демографической группы в летописи окаменелостей человека. Частота встречаемости более 45 человек постепенно увеличивается со временем, но в целом оставалась довольно низкой на протяжении всего палеолита. У него также были возрастные недуги в пояснице. кифоз (чрезмерное искривление поясничных позвонков нижней части спины), L5 – S1 спондилолистез (несовпадение последнего поясничного позвонка с первым крестцовый позвонок ), и Болезнь Бааструпа на L4 и 5 (увеличение остистых отростков). Это вызвало бы боль в пояснице, значительно ограничивало бы движения и могло свидетельствовать о групповом уходе.[33]
У подросткового черепа SH (Cranium 14) диагностирован ламбдовидный краниосиностоз с одинарным швом (незрелое закрытие левой ламбдовидный шов, что привело к деформации черепа по мере продолжения развития). Это редкое заболевание, встречающееся менее чем у 6 из каждых 200 000 человек современного человека. Этот человек умер в возрасте около 5 лет, что позволяет предположить, что от него не отказались из-за его деформации, как это делалось в исторические времена, и он получил такое же качество ухода, как и любой другой ребенок.[34]
Гипоплазия эмали на зубах используется для определения приступов пищевого стресса. На 40% это значительно выше, чем в более ранних южноафриканских гоминин Paranthropus robustus в Сварткранс (30,6%) или Стеркфонтейн (12,1%). Тем не менее, неандертальцы страдали еще более высокими показателями и более интенсивными приступами гипоплазии, но неясно, связано ли это с тем, что неандертальцы были менее способны к эксплуатации природных ресурсов или жили в более суровых условиях. Пик в возрасте 3,5 лет может коррелировать с возрастом отлучения от груди. У неандертальцев этот пик пришелся на 4 года, и многие современные охотники-собиратели также отлучают от груди примерно в 4 года.[35]
Культура
Еда

Сообщества среднего плейстоцена в целом, по-видимому, ели крупную дичь чаще, чем их предшественники, при этом мясо становится важным диетическим компонентом. Диета в целом может быть разнообразной - например, жители Терра Аматы, похоже, ели в основном оленей, но также слонов, кабанов, горных козлов, носорогов и других животных. зубр. На африканских участках обычно встречаются кости крупного рогатого скота и лошади. Хотя туши могли быть просто вычищены, некоторые афро-европейские участки демонстрируют особую нацеленность на один вид, что более вероятно указывает на активную охоту; Например: Olorgesailie, Кения, где было добыто от 50 до 60 особей павианов (Theropithecus oswaldi ); а также Торральба и Амброна в Испании, в которых много слоновьих костей (а также носорогов и крупных копытных млекопитающих). Увеличение потребления мяса может указывать на развитие стратегии групповой охоты в среднем плейстоцене. Например, в Торральбе и Амброне животные могли попасть в болота, прежде чем были убиты, что повлекло за собой окружение и движение большой группой охотников в рамках скоординированной и организованной атаки. Эксплуатация водной среды, как правило, отсутствует, несмотря на то, что некоторые участки находятся в непосредственной близости от океана, озер или рек.[36]
Растения, вероятно, также часто употреблялись в пищу, в том числе сезонно доступные, но степень их использования неясна, поскольку они не окаменели так же, как кости животных. Если предположить, что диета состоит из нежирного мяса, человеку потребуется углевод потребление, чтобы предотвратить белковое отравление, например, обычно есть много под землей органы хранения, кора деревьев, ягоды или орехи. В Schöningen в окрестностях Германии насчитывается более 200 растений, которые можно съедать в сыром или приготовленном виде.[37]
Изобразительное искусство

Современные люди верхнего палеолита хорошо известны гравировками, имеющими, по-видимому, символическую ценность. По состоянию на 2018 год только 27 объектов среднего и нижнего палеолита были постулированы как имеющие символическую гравировку по состоянию на 2018 год, из которых некоторые были опровергнуты как вызванные природными или иными несимволическими явлениями (такими как процессы окаменелости или раскопок). . К нижнепалеолитическим относятся: три гальки возрастом 380 000 лет из Терра Аматы; галька возрастом 250 000 лет из Марклеберг, Германия; 18 камешков возрастом примерно 200000 лет из Лазарет (около Терра Амата); каменный камень возрастом около 200000 лет из Grotte de l'Observatoire, Монако; кость возрастом 370000 лет из Bilzingsleben, Германия; и галька возрастом от 200 до 130 тысяч лет из Baume Bonne, Франция.[38]
В середине 19 века французский археолог Жак Буше де Кревекёр де Перт начали раскопки в Св. Ачеуле, Амьен, Франция, (район, где был определен ашельский ярус), и, помимо ручных топоров, сообщалось об окаменелостях перфорированной губки (Porosphaera globularis ), которые он считал декоративными бусинами. Это утверждение было полностью проигнорировано, и Буше де Перт в ответ на академическое молчание сказал: «[Археологи] применили против меня оружие более мощное, чем возражения, чем критика, сатира или даже преследование - оружие презрения. не обсуждали мои факты, они даже не потрудились отрицать их. Они проигнорировали их ". В 1894 г. английский археолог Уортингтон Джордж Смит обнаружил 200 подобных перфорированных окаменелостей в Бедфордшир, Англия, а также предполагали, что они выполняют функцию бусинок, хотя не упомянули находку Буше де Пертеса, возможно, потому, что он не знал об этом. В 2005 году Роберт Беднарик пересмотрел материал и пришел к выводу, что - потому что весь Бедфордширский P. globularis окаменелости имеют субсферическую форму и колеблются в диапазоне 10–18 мм (0,39–0,71 дюйма) в диаметре, несмотря на то, что этот вид имеет очень изменчивую форму - они были выбраны сознательно. Похоже, они были полностью или почти полностью проданы каким-то паразитическим существом (т.е., в результате естественных процессов), а затем их ударили по тому, что было бы более закрытым концом, чтобы полностью открыть отверстие. Он также обнаружил изношенные грани, которые, как он предполагал, возникли в результате стука о другие бусины, когда их связывали вместе и носили как ожерелье.[39] В 2009 году Соланж Риго, Франсиско д'Эррико и коллеги заметили, что измененные области светлее по цвету, чем немодифицированные, предполагая, что они были нанесены гораздо позже, например, во время раскопок. Они также не были убеждены в том, что окаменелости могут быть достоверно связаны с ашельскими артефактами из этих мест, и также предположили, что - в качестве альтернативы архаической деятельности человека - очевидный выбор размера мог быть вызван либо естественными геологическими процессами, либо предпочтениями коллекционеров XIX века. эта конкретная форма.[40]
Ранние современные люди и поздние неандертальцы (последние особенно после 60 000 лет назад) широко использовали красный цвет. охра предположительно для символических целей, поскольку она имеет цвет, похожий на кровь, хотя охра также может иметь функциональное лекарственное применение. За пределами этих двух видов охра употребляется в Олдувайское ущелье, Танзания, где были обнаружены два куска красной охры; Амброна, где плита охры была обрезана до определенной формы; и Terra Amata, где 75 кусочков охры были нагреты для получения широкого диапазона цветов от желтого до красно-коричневого и красного. Они могут служить примером ранних и единичных случаев цветового предпочтения и цветовой катагоризации, и такие практики, возможно, еще не были нормализованы.[41]

В 2006 году Эудальд Карбонелл и Марина Москера предположили, что гоминины SH были похоронены людьми, а не жертвами какого-то катастрофического события, такого как пещера, потому что маленькие дети и младенцы отсутствуют, что было бы неожиданно, если бы это была одна и полная семья. единица. Люди SH явно ассоциируются только с одним каменным инструментом, тщательно обработанным ручным топором высокого качества (и редко используемым в регионе). кварцит, поэтому Карбонелл и Москера постулировали, что это было намеренно и символически помещено вместе с телами как некое подобие могилы. Предполагаемые доказательства символических могил не обнаружатся еще 300000 лет.[42]
Технологии
Каменные инструменты
Нижний палеолит (ранний каменный век) включает в себя Oldowan который был заменен Ашельский характеризуется именно производством в основном симметричных ручные топоры. У ашельцев период времени составляет около миллиона лет, и такая технологическая стагнация обычно объясняется сравнительно ограниченными когнитивными способностями, которые значительно снижают инновационный потенциал, например дефицитом когнитивной текучести, рабочая память, или социальная система, совместимая с ученичеством. Тем не менее, ашельский период, кажется, слегка меняется со временем и обычно делится на ранний ашельский и поздний ашельский, причем последний стал особенно популярным после 600-500 тысяч лет назад. Позднеахельские технологии никогда не переходили к востоку от Movius Line в Восточную Азию, что, как обычно считается, связано с серьезным дефицитом культурной передачи, а именно с меньшим размером населения на Востоке.[43]

На переход указывает производство меньших, более тонких и более симметричных ручных топоров (хотя все еще производились более толстые и менее изысканные топоры). В возрасте 500000 лет Boxgrove участок, Англия - исключительно хорошо сохранившийся участок с обилием остатков инструментов. Прореживание могло быть произведено путем ударов ручного топора почти перпендикулярно мягким молотком, что возможно с изобретением подготовленных платформ для изготовления инструментов. Срезатели Boxgrove также оставили после себя большие каменные хлопья остатки от изготовления ручных топоров, возможно, с намерением переработать их в какой-нибудь другой инструмент позже. Позднеахельские памятники в других местах заранее подготовлены каменные ядра ("Large Flake Blanks", LFB) различными способами, прежде чем преобразовывать их в инструменты, делая подготовленные платформы ненужными. LFB Acheulian распространился из Африки в Западную и Южную Азию до миллиона лет назад и присутствует в Южной Европе после 600000 лет назад, но северная Европа (и Левант после 700000 лет назад) использовали мягкие молотки, поскольку они в основном делали нас из маленьких, толстых кремень узелки. Первые подготовленные платформы в Африке относятся к возрасту 450 000 лет. Форезмит промышленность, переходное между Ранний каменный век (Ашельский) и Средний каменный век.[43]
При использовании любого из этих методов разработчикам инструментов (производителям инструментов) пришлось бы производить какой-то элемент, косвенно связанный с созданием желаемого продукта (иерархическая организация), который мог бы представлять собой серьезное когнитивное развитие. Эксперименты с современными людьми показали, что подготовка платформы не может быть изучена с помощью чисто наблюдательного обучения, в отличие от более ранних методов, и может быть признаком хорошо разработанных методов обучения, а также саморегулируемое обучение. В Боксгроуве мастерицы (изготовители инструментов) использовали не только камень, но также кости и рога для изготовления молотки, и использование такого широкого диапазона сырья может говорить о расширенных возможностях планирования, поскольку обработка камня требует совершенно другого набора навыков для работы и сбора материалов, чем для работы с костями.[43]
В Каптуринская свита В Кении, были обнаружены самые старые свидетельства технологии лезвий и лезвий, датируемые 545–509 тысячами лет назад. Эта технология редко встречается даже в среднем палеолите и обычно связана с Верхний палеолит современные люди. Неясно, является ли это частью давней традиции изготовления клинков, или технология клинков была утеряна и изобретена несколько раз множеством разных человеческих видов.[44]
Пожар и строительство
Несмотря на очевидные толчки в более холодный климат, свидетельства пожара в археологических записях немногочисленны и редки, вплоть до 400–300 тысяч лет назад. Хотя возможно, что остатки пожара просто деградировали, длинные и в целом ненарушенные последовательности оккупации, такие как в Араго или Гран-Долина, явно не имеют убедительных доказательств использования огня. Этот образец, возможно, может указывать на изобретение технологии зажигания или улучшенных методов обслуживания пожара в то время, и что огонь не был неотъемлемой частью жизни людей до этого в Европе. В Африке, с другой стороны, люди, возможно, уже 1,6 миллиона лет назад могли часто убирать огонь в результате естественных лесных пожаров, которые происходят гораздо чаще на континенте, таким образом, возможно (более или менее) регулярно используя огонь. Самому старому установленному непрерывному пожару за пределами Африки 780 000 лет. Гешер Бенот Яаков, Израиль.[45]
В Европе свидетельства построенных жилых конструкций - классифицируемых как хижины с твердой поверхностью и прочным фундаментом, построенные в районах, в основном защищенных от непогоды, - были зарегистрированы с момента Кромеровское межледниковье, самый ранний пример - каменный фундамент возрастом 700000 лет из Пржезлетице, Чехия. Это жилище, вероятно, имело сводчатую крышу из толстых ветвей или тонких столбов, опирающуюся на фундамент из больших камней и земли. Было высказано предположение, что другие такие жилища существовали во время или после Гольштейн Межледниковье (который начался 424 000 лет назад) в Бильзингслебене, Германия; Terra Amata, Франция; и Fermanville и Сен-Жермен-де-Во в Нормандия. Вероятно, они были заняты зимой, и, имея в среднем площадь 3,5 м × 3 м (11,5 футов × 9,8 футов), они, вероятно, использовались только для сна, в то время как другие действия (включая пожаротушение), похоже, выполнялись на улице. Технология менее прочных палаток могла быть изобретена в Европе в нижнем палеолите.[46]
Спирс

Возникновение повторного использования огня - самое раннее в Европе из Бич-Пита, Англия и Шенинген, Германия - примерно совпадает с рывок технология (прикрепление каменных наконечников к копьям) лучше всего иллюстрируется Копья Шёнингена.[45] Эти 9 деревянных копий и фрагментов копий - в дополнение к копье и двухконечной палке - датируются 300 000 лет назад и хранились на берегу озера. Копья варьируются от 2,9–4,7 см (1,1–1,9 дюйма) в диаметре и, возможно, были длиной 210–240 см (7–8 футов), что в целом похоже на современные соревновательные копья. Копья были сделаны из мягких ель дерева, за исключением копья 4, которое было (также мягким) сосна дерево. Это контрастирует с Клактон наконечник копья из Клактон-он-Си, Англия, возможно, примерно на 100 000 лет старше, сделанная из твердого тис дерево. Копья Шенингена могли иметь дальность действия до 35 м (115 футов), хотя были бы более эффективными на коротких дистанциях в пределах 5 м (16 футов), что делало их эффективным оружием дальнего боя как против добычи, так и против хищников. Помимо этих двух мест, единственное другое место, которое предоставляет убедительные доказательства европейской технологии копья, - это здание, которому 120000 лет. Lehringen место, Германия, где тисовое копье диаметром 238 см (8 футов), по-видимому, было воткнуто в слона.[47] В Африке 500000-летние точки от Кату Пан 1, Южная Африка, возможно, были надеты на копья. Судя по косвенным данным, лошадь лопатка в Боксгроуве, которому 500000 лет, показана колотая рана, похожая на рану от копья. Признаки хафтинга (как в Европе, так и в Африке) становятся все более распространенными в Средний каменный век.[48]
Язык
У людей SH был современный человекоподобный подъязычная кость (который поддерживает язык), и среднее ухо кости, способные точно различать частоты в диапазоне нормальной человеческой речи. Судя по зубным бороздкам, они, по-видимому, были преимущественно праворукими, а их праворукость связана с латерализация функции мозга, обычно связанный с обработкой языка у современных людей. Итак, предполагается, что это население разговаривало с некоторыми ранняя форма языка.[42][49][50] Тем не менее, эти черты не являются абсолютно доказательством существования языка и речи, подобной человеку, и ее присутствие на столь раннем этапе, несмотря на такие анатомические аргументы, в первую очередь отвергалось учеными-когнитивистами Филип Либерман.[49]
Смотрите также
Рекомендации
- ^ Харвати, К. (2007). «100 лет Homo heidelbergensis - жизнь и времена спорного таксона» (PDF). Mitteilungen der Gesellschaft für Urgeschichte. 16: 85. Архивировано с оригинал (PDF) 4 марта 2016 г.
- ^ а б c Mounier, A .; Marchal, F .; Кондеми, С. (2009). "Является Homo heidelbergensis отдельный вид? Новое понимание нижней челюсти Мауэра ". Журнал эволюции человека. 56 (3): 219–46. Дои:10.1016 / j.jhevol.2008.12.006. PMID 19249816.
- ^ Бегун, Дэвид Р., изд. (2012). "Африканское происхождение Homo sapiens". Спутник палеоантропологии. Джон Вили и сыновья. ISBN 9781118332375.
- ^ а б Шварц, Дж. Х.; Таттерсолл, И. (2010). "Ископаемые свидетельства происхождения Homo sapiens". Американский журнал физической антропологии. 143 (S51): 96–98, 101–103. Дои:10.1002 / ajpa.21443. PMID 21086529.
- ^ а б Стрингер, К. (2012). "Статус Homo heidelbergensis (Schoetensack 1908) ". Эволюционная антропология. 21 (3): 101–104. Дои:10.1002 / evan.21311. PMID 22718477. S2CID 205826399.
- ^ а б де Ламли, М.-А. (2015). "L'homme de Tautavel. Un человек прямоходящий européen évolué. Homo erectus tautavelensis". L'Anthropologie. 119 (3): 344–346. Дои:10.1016 / j.anthro.2015.06.001.
- ^ а б c Buck, L.T .; Стрингер, К. (2014). "Homo heidelbergensis". Текущая биология. 24 (6): R214 – R215. Дои:10.1016 / j.cub.2013.12.048. ISSN 0960-9822. PMID 24650901.
- ^ "Hierzu zählte noch im Jahr 2010 auch das Geologisch-Paläontologische Institut der Universität Heidelberg, das den Unterkiefer seit 1908 verwahrt und ihn als Homo erectus heidelbergensis auswies. Inzwischen wird er jedoch auch in Heidelberg als Homo heidelbergensis bezeichnet, siehe " (на немецком). Sammlung des Instituts für Geowissenschaften. Получено 29 ноября, 2015.
- ^ de Castro, J.M.B .; Мартинес, И .; Gracia-Téllez, A .; Martinón ‐ Torres, M .; Арсуага, Дж. Л. (2020). «Стоянка гомининов среднего плейстоцена Сима-де-лос-Уэсос (Бургос, Испания). Оценка количества особей». Анатомический рекорд. Дои:10.1002 / ar.24551.
- ^ Мейер, М .; Арсуага, Ж.-Л.; de Filippo, C .; и другие. (2016). "Последовательности ядерной ДНК из гомининов Sima de los Huesos среднего плейстоцена". Природа. 531: 504–507. Дои:10.1038 / природа17405.
- ^ Стрингер, Крис (2012). «Комментарий: Что делает современного человека». Природа. 485 (7396): 33–35 [34]. Bibcode:2012Натура 485 ... 33С. Дои:10.1038 / 485033a. PMID 22552077. S2CID 4420496.
- ^ Грюн, Райнер; Пайк, Алистер; Макдермотт, Фрэнк; Эггинс, Стивен; Мортимер, Грэм; Обер, Максим; Кинсли, Лесли; Жоанн-Бояу, Рено; Рамси, Майкл; Денис, Кристиана; Бринк, Джеймс; Кларк, Тара; Стрингер, Крис (1 апреля 2020 г.). «Датировка черепа из Брокен-Хилла, Замбия, и его положение в эволюции человека». Природа. 580 (7803): 372–375. Bibcode:2020Натура.580..372Г. Дои:10.1038 / с41586-020-2165-4. PMID 32296179. S2CID 214736650.
- ^ Белый, Тим Д.; Asfaw, B .; DeGusta, D .; Gilbert, H .; Richards, G.D .; Suwa, G .; Хауэлл, Ф. К. (2003). «Плейстоцен Homo sapiens из Среднего Аваша, Эфиопия ". Природа. 423 (6491): 742–747. Bibcode:2003 Натур.423..742Вт. Дои:10.1038 / природа01669. PMID 12802332. S2CID 4432091.
- ^ Асфау, Берхане (2005). «Новая теменная часть гоминида из Бодо, средняя долина Аваш, Эфиопия». Американский журнал физической антропологии. 61 (3): 367–371. Дои:10.1002 / ajpa.1330610311. PMID 6412559.
- ^ Райтмайр, Дж. Филип (2005). "Череп озера Ндуту и ранний Homo sapiens в Африке". Американский журнал физической антропологии. 61 (2): 245–254. Дои:10.1002 / ajpa.1330610214. PMID 6410925.
- ^ а б c Профико, А .; ди Винченцо, Ф .; и другие. (2016). "Заполнение пробела. Человеческие черепные останки из Гомборе II (Мелка Кунтура, Эфиопия; ок. 850 тыс. Лет назад) и происхождение Homo heidelbergensis". Журнал антропологических наук. 94: 1–24. Дои:10.4436 / JASS.94019.
- ^ de Castro, J.M.B .; Martinón ‐ Torres, M .; Арсуага, Дж. Л.; Карбонелл, Э. (2017). "Двадцатилетие Гомо-предшественник (1997-2017): обзор ». Эволюционная антропология. 26 (4): 164. Дои:10.1002 / evan.21540.
- ^ Райтмайр, Дж. Филип (1998). «Эволюция человека в среднем плейстоцене: роль Homo heidelbergensis" (PDF). Эволюционная антропология. 6 (6): 218–227. Дои:10.1002 / (SICI) 1520-6505 (1998) 6: 6 <218 :: AID-EVAN4> 3.0.CO; 2-6.
- ^ Мейер, М .; Arsuaga, J .; de Filippo, C .; Нагель, С. (2016). "Последовательности ядерной ДНК из гомининов Sima de los Huesos среднего плейстоцена". Природа. 531 (7595): 504–507. Bibcode:2016Натура.531..504M. Дои:10.1038 / природа17405. PMID 26976447. S2CID 4467094.
- ^ Bermudez de Castro, J.M .; Арсуага, Дж. Л.; Carbonell, E .; Rosas, A .; Мартинес, И .; Москера, М. (1997). «Гоминид из нижнего плейстоцена Атапуэрка, Испания: возможный предок неандертальцев и современных людей» (PDF). Наука. 276 (5317): 1392–1395. Дои:10.1126 / science.276.5317.1392. PMID 9162001. S2CID 31088294.
- ^ Welker, F .; Ramos-Madrigal, J .; Gutenbrunner, P .; и другие. (2020-04-01). «Зубной протеом Homo antecessor». Природа. 580 (7802): 235–238. Bibcode:2020Natur.580..235W. Дои:10.1038 / с41586-020-2153-8. ISSN 1476-4687. PMID 32269345. S2CID 214736611.
- ^ Hosfield, R .; Коул, Дж. (2018). «Ранние гоминины в северо-западной Европе: длинная пунктирная хронология?». Четвертичные научные обзоры. 190: 148–160. Дои:10.1016 / j.quascirev.2018.04.026.
- ^ а б Райтмайр, Р. П. (2004). "Размер мозга и энцефализация в период от раннего до среднего плейстоцена. Гомо". Американский журнал физической антропологии. 124 (2): 109–123. Дои:10.1002 / ajpa.10346. PMID 15160365.
- ^ Allen, J. S .; Damasio, H .; Грабовски, Т. Дж. (2002). «Нормальные нейроанатомические изменения в мозге человека: объемное МРТ-исследование». Американский журнал физической антропологии. 118 (4): 341–358. Дои:10.1002 / ajpa.10092. PMID 12124914. S2CID 21705705.
- ^ Стрингер, К. (1984). «Эволюция человека и биологическая адаптация в плейстоцене». В Фоли, Р. (ред.). Эволюция гоминидов и экология сообщества. Академическая пресса. ISBN 978-0-12-261920-5.
- ^ Холлоуэй, Р. Л. (1985). "Бедный мозг Homo sapiens neanderthalensis: смотрите, что вам нравится ". В Делсон, Э. (ред.). Предки: веские доказательства. Алан Р. Лисс. ISBN 978-0-471-84376-4.
- ^ Amano, H .; Kikuchi, T .; Morita, Y .; Кондо, О .; Suzuki, H .; и другие. (2015). "Виртуальная реконструкция черепа неандертальца Амуд 1" (PDF). Американский журнал физической антропологии. 158 (2): 185–197. Дои:10.1002 / ajpa.22777. HDL:10261/123419. PMID 26249757.
- ^ Карретеро, Хосе-Мигель; Родригес, Лаура; Гарсиа-Гонсалес, Ребека; Арсуага, Хуан-Луис; Гомес-Оливенсия, Азиер; Лоренцо, Карлос; Бонмати, Алехандро; Грация, Ана; Мартинес, Игнасио (2012). «Оценка роста длинных костей у людей среднего плейстоцена из Сима-де-лос-Уэсос, Сьерра-де-Атапуэрка (Испания)» (PDF). Журнал эволюции человека. 62 (2): 242–55. Дои:10.1016 / j.jhevol.2011.11.004. PMID 22196156.
- ^ а б Арсуага, Дж. Л.; Carretero, J.-M .; Lorenzo, C .; и другие. (2015). «Посткраниальная морфология людей среднего плейстоцена из Сима-де-лос-Уэсос, Испания». PNAS. 112 (37): 11524–11529. Дои:10.1073 / pnas.1514828112.
- ^ ДеКроос, ФК; Liao, JC; Ramey, NA; Ли, я (2011-08-15). «Лечение одонтогенного орбитального целлюлита». Журнал медицины и жизни. 4 (3): 314–317. ISSN 1844–122X. ЧВК 3168817. PMID 22567060.
- ^ Ascaso, F .; Адиего, М. (2016-09-14). «Homo heidelbergensis: самый старый случай одонтогенного целлюлита орбиты?». Acta Ophthalmologica. 94. Дои:10.1111 / j.1755-3768.2016.0022. ISSN 1755–375X.
- ^ Грасиа-Телес, Ана; Арсуага, Хуан-Луис; Мартинес, Игнасио; Мартин-Франсес, Лаура; Мартинон-Торрес, Мария; Бермудес де Кастро, Хосе-Мария; Бонмати, Алехандро; Лира, Хайме (2013-05-08). «Орофациальная патология у Homo heidelbergensis: Случай черепа 5 из Сайма-де-лос-Уэсос (Атапуэрка, Испания)». Четвертичный международный. 295: 83–93. Bibcode:2013 г., 295 ... 83 г. Дои:10.1016 / j.quaint.2012.02.005. ISSN 1040-6182.
- ^ Bonmatí, A .; Gómez-Olivencia, A .; Арсуага, Дж. Л.; и другие. (2010). «Нижняя часть спины и таз среднего плейстоцена от пожилого человека из поселения Сима-де-лос-Уэсос, Испания». Труды Национальной академии наук. 107 (43): 18386–18391. Дои:10.1073 / pnas.1012131107. ЧВК 2973007. PMID 20937858.
- ^ Garcia, A .; Арсуага, Дж. Л.; Мартинес, И .; и другие. (2009). "Краниосиностоз в среднем плейстоцене череп 14 человека из Сима-де-лос-Уэсос, Атапуэрка, Испания". Дои:10.1073 / pnas.0900965106. ЧВК 2672549. PMID 19332773. Цитировать журнал требует
| журнал =(помощь) - ^ Bermúdez de Castro, J.M .; Перес, П. Дж. (1995). «Гипоплазия эмали гоминидов среднего плейстоцена из Атапуэрка (Испания)». Американский журнал физической антропологии. 96: 301–314. Дои:10.1002 / ajpa.1330960307.
- ^ Исаак, Г. (1971). «Рацион древнего человека: аспекты археологических раскопок из памятников нижнего и среднего плейстоцена в Африке». Мировая археология. 2 (3): 279–299. Дои:10.1080/00438243.1971.9979481.
- ^ Bigga, G .; Schoch, W. H .; Урбан, Б. (2015). «Палеосреда и возможности эксплуатации растений в среднем плейстоцене Шёнингена (Германия). Выводы из ботанических макро останков и пыльцы». Журнал эволюции человека. 89: 92–104. Дои:10.1016 / j.jhevol.2015.10.005.
- ^ Majkić, A .; d’Errico, F .; Степанчук, В. (2018). «Оценка значимости коры с гравировкой палеолита. Пример из мустьерского городища Киик-Коба, Крым». PLOS ONE. 13 (5): e0195049. Bibcode:2018PLoSO..1395049M. Дои:10.1371 / journal.pone.0195049. ЧВК 5931501. PMID 29718916.
- ^ Беднарик, Р. Г. (2005). "Подробнее об ашельском бисере". Исследования наскального искусства. 22 (2): 210–212.
- ^ Rigaud, S .; d'Errico, F .; Vanhaeren, M .; Нойман, К. (2009). "Критическая переоценка предполагаемого ашельского Porosphaera globularis бусы ». Журнал археологической науки. 36 (1): 25–34.
- ^ Wreschner, E. E .; Bolton, R .; и другие. (1980). «Красная охра и эволюция человека: повод для обсуждения» (PDF). Современная антропология. 21 (5): 631–632. Дои:10.1086/202541. JSTOR 2741829.
- ^ а б c Carbonell, E .; Москера, М. (2006). «Возникновение символического поведения: могильная яма Сима-де-лос-Уэсос, Сьерра-де-Атапуэрка, Бургос, Испания». Comptes Rendus Palevol. 5 (1–2): 155–160. Дои:10.1016 / j.crpv.2005.11.010.
- ^ а б c Стаут, Д .; Apel, J .; Commander, J .; Робертс, М. (2014). «Поздние ашельские технологии и познание в Boxgrove, Великобритания». Журнал археологической науки. 41: 576–590.
- ^ Johnson, C. R .; МакБриарти, С. (2010). «Лезвия возрастом 500 000 лет из формации Каптурин, Кения». Журнал эволюции человека. 58 (2): 193–200. Дои:10.1016 / j.jhevol.2009.10.001.
- ^ а б Roebroekes, W .; Вилла, П. (2011). «О самых ранних свидетельствах использования огня в Европе». Труды Национальной академии наук. 108 (13): 5209–5214. Bibcode:2011ПНАС..108.5209Р. Дои:10.1073 / pnas.1018116108. ЧВК 3069174. PMID 21402905.
- ^ Скленарж, К. (1987). «Жилая структура нижнего палеолита в Пржезлетицах и ее значение». Антропология. 25 (2): 101–103. JSTOR 26294864.
- ^ Schoch, W. H .; Bigga, G .; Böhner, U .; Richter, P .; Тербергер, Т. (2015). «Новые сведения о деревянном оружии из палеолитической стоянки Шенинген». Журнал эволюции человека. 89: 214–225. Дои:10.1016 / j.jhevol.2015.08.004.
- ^ Уилкинс, Джейн; Schoville, Benjamin J .; Браун, Кайл С .; Чазан, Майкл (2012). "Свидетельства ранней технологии охоты" (PDF). Наука. 338 (6109): 942–6. Bibcode:2012Sci ... 338..942W. Дои:10.1126 / science.1227608. PMID 23161998. S2CID 206544031.
- ^ а б Мартинес, И .; Арсуага, Дж. Л.; Quam, R .; и другие. (2008). "Подъязычные кости человека из среднего плейстоцена участка Сима-де-лос-Уэсос (Сьерра-де-Атапуэрка, Испания)". Журнал эволюции человека. 54 (1): 118–124.
- ^ Лозано, Марина; Москера, Марина; Де Кастро, Хосе Мария Бермудес; Арсуага, Хуан Луис; Карбонелл, Эудальд (2009). "Праворукость Homo heidelbergensis из Сима-де-лос-Уэсос (Атапуэрка, Испания) 500 000 лет назад ". Эволюция и поведение человека. 30 (5): 369–76. Дои:10.1016 / j.evolhumbehav.2009.03.001.
внешняя ссылка
- Homo heidelbergensis - Программа Смитсоновского института по происхождению человека
- Домашняя страница Mauer 1 Club
- Центр всемирного наследия ЮНЕСКО - Археологические раскопки Атапуэрка
- Хронология человека (интерактивная) – Смитсоновский институт, Национальный музей естественной истории (Август 2016 г.).
| Археология |
| ||||
|---|---|---|---|---|---|
| Переходные окаменелости с Х. heidelbergensis черты |
| ||||
| Родственные виды | |||||
| |||||
| Таксономия (Гоминины ) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Предки |
| ||||||||||||||||||||||||||
| Модели |
| ||||||||||||||||||||||||||
| Сроки | |||||||||||||||||||||||||||
| Другие |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| Авторитетный контроль |
|---|