Субталамическое ядро - Subthalamic nucleus
| Субталамическое ядро | |
|---|---|
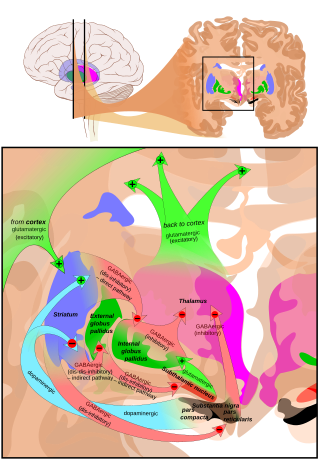
 Корональные срезы человеческого мозга, показывающие базальный ганглий (внешний бледный шар (GPe) и внутренний бледный шар (GPi)), субталамическое ядро (STN) и черная субстанция (SN). | |
 DA-петли в болезнь Паркинсона | |
| Подробности | |
| Часть | Базальный ганглий |
| Идентификаторы | |
| латинский | ядро subthalamicus |
| MeSH | D020531 |
| NeuroNames | 435 |
| НейроЛекс Я БЫ | nlx_anat_1010002 |
| TA98 | A14.1.08.702 |
| TA2 | 5709 |
| FMA | 62035 |
| Анатомические термины нейроанатомии | |
В субталамическое ядро это маленькая линза в форме ядро в мозг где он, с функциональной точки зрения, является частью базальный ганглий система. С точки зрения анатомии, это основная часть субталамус. Как следует из названия, субталамическое ядро расположено вентральный к таламус. Это также дорсально по отношению к черная субстанция и медиальнее внутренняя капсула. Впервые он был описан Жюль Бернар Луйс в 1865 г.,[1] и срок корпус Луйси или же Тело Луйса до сих пор иногда используется.
Анатомия

Структура
Основной вид нейрон находится в субталамическом ядре, имеет довольно длинный, редко колючий дендриты.[2][3] В более центрально расположенных нейронах дендритные ветви имеют больше эллипсоидальный форма.[4] Размеры этих беседок (1200 мкм, 600 мкм и 300 мкм) одинаковы для многих видов, включая крыс, кошек, обезьян и человека, что необычно. Однако количество нейронов увеличивается с увеличением размера мозга, а также с увеличением внешних размеров ядра. Основные нейроны: глутаматергический, которые придают им особое функциональное положение в системе базальных ганглиев. У людей также имеется небольшое количество (около 7,5%) ГАМКергический интернейроны которые участвуют в местных схемах; однако дендритные ветви субталамических нейронов уклоняются от границы и в первую очередь взаимодействуют друг с другом.[5]
Афферентные аксоны
Субталамическое ядро получает основной вклад от внешний бледный шар (GPe),[6] не столько через ansa lenticularis как часто говорят, но излучающие волокна сначала пересекают медиальный pallidum и внутреннюю капсулу (см. рисунок). Эти афференты являются ГАМКергическими, угнетающими нейроны в субталамическом ядре. Возбуждающие глутаматергические входы исходят от кора головного мозга (в частности, моторная кора), а из частей парафасцикулярная мышца из центральный комплекс. Субталамическое ядро также получает нейромодулирующий входы, в частности дофаминергический аксоны из черная субстанция pars compacta.[7]Он также получает входные данные от педункулопонтинное ядро.
Эфферентные цели
Аксоны нейронов субталамического ядра покидают ядро дорсально. Эфферентные аксоны глутаматергические (возбуждающие). За исключением соединения со стриатумом (17,3% у макак), большинство основных нейронов субталамической области являются многоцелевыми и направлены на другие элементы ядра базальных ганглиев.[8] Некоторые направляют аксоны к черной субстанции медиально и к медиальному и латеральному ядрам паллидума латерально (3 мишени, 21,3%). Некоторые из них являются двумя мишенями с латеральным паллидумом и черной субстанцией (2,7%) или латеральным паллидумом и медиальной частью (48%). Реже - единственная цель для латерального паллидума. В паллидуме субталамические окончания заканчиваются полосами, параллельными паллидальной границе.[8][9] Когда все аксоны, достигающие этой мишени, добавляются, основное влияние субталамического ядра в 82,7% случаев очевидно внутренний бледный шар (ГПи).
Некоторые исследователи сообщили о внутренних аксон залоги.[10] Однако функциональных доказательств этому мало.
Физиология
Субталамическое ядро
Первые внутриклеточные электрические записи субталамических нейронов были выполнены с использованием острых электродов в препарате крысиного среза.[нужна цитата ] В этих записях были сделаны три ключевых наблюдения, все три из которых преобладали в последующих отчетах о субталамических свойствах воспламенения. Первое наблюдение заключалось в том, что в отсутствие инъекции тока или синаптической стимуляции большинство клеток запускались спонтанно. Второе наблюдение заключается в том, что эти клетки способны временно срабатывать на очень высоких частотах. Третье наблюдение касается нелинейного поведения, когда клетки временно деполяризованы после гиперполяризации ниже -65 мВ. Затем они могут задействовать управляемые по напряжению кальциевые и натриевые токи, чтобы вызвать всплески потенциалов действия.
Несколько недавних исследований были сосредоточены на способности субталамических нейронов управлять автономным ритмом. Эти клетки часто называют «кардиостимуляторами с быстрым импульсом»,[11] поскольку они могут генерировать спонтанные потенциалы действия с частотой от 80 до 90 Гц у приматов.
Колебательная и синхронная деятельность[12][13] вероятно, это типичный образец разряда в субталамических нейронах, зарегистрированный у пациентов и животных моделей, характеризующийся потерей дофаминергических клеток в черная субстанция pars compacta, что является основной патологией, лежащей в основе болезнь Паркинсона.
Латеропаллидо-субталамическая система
Сильные реципрокные связи связывают субталамическое ядро и внешний сегмент бледный шар. Оба являются кардиостимуляторами с высокой скоростью. Считается, что вместе они составляют «центральный кардиостимулятор базальных ганглиев».[14] с синхронными всплесками.
Соединение латерального паллидума с субталамическим ядром также является соединением в базальный ганглий система, в которой сокращение между передающими / приемными элементами, вероятно, является самым сильным. Что касается объема, у людей размер латерального паллидума составляет 808 мм³, а субталамического ядра - всего 158 мм³.[15] Это выражается в количестве нейронов, что представляет собой сильное сжатие с потерей точности карты.
Некоторые аксоны от латерального pallidum переходят в полосатое тело.[16] На активность медиального паллидума влияют афферентность латерального паллидума и субталамического ядра.[17] То же самое для черная субстанция pars reticulata.[9] Субталамическое ядро отправляет аксоны другому регулятору: педункуло-понтинному комплексу (id).
Считается, что латеропаллидо-субталамическая система играет ключевую роль в формировании паттернов активности, наблюдаемых в болезнь Паркинсона.[18]
Патофизиология
Хроническая стимуляция СТН, называемая глубокая стимуляция мозга (DBS), используется для лечения пациентов с болезнь Паркинсона. В первую очередь стимулируются терминальные разветвления афферентных аксонов, которые изменяют активность субталамических нейронов. Однако на срезах таламуса мышей было показано, что[19] что стимул также вызывает высвобождение ближайших астроцитов аденозинтрифосфат (АТФ), предшественник аденозин (через катаболический процесс). В свою очередь, активация аденозинового рецептора A1 подавляет передачу возбуждения в таламусе, имитируя тем самым абляция субталамического ядра.
Одностороннее разрушение или разрушение субталамического ядра, которое обычно может происходить в результате инсульта небольшого сосуда у пациентов с диабетом, гипертонией или курением в анамнезе, вызывает гемибаллизм.
Поскольку одна из предполагаемых функций STN заключается в управлении импульсами, дисфункция в этой области была связана с обсессивно-компульсивное расстройство.[20] Искусственная стимуляция STN показала некоторые перспективы в коррекции тяжелого импульсивного поведения и впоследствии может быть использована в качестве альтернативного лечения расстройства.[21]
Функция
Функция STN неизвестна, но современные теории помещают ее как компонент базальный ганглий система управления, которая может выполнять выбор действия. Считается, что он реализует так называемый «гиперпрямой путь» моторного контроля, в отличие от прямых и непрямых путей, реализуемых где-то еще в базальных ганглиях. Также было показано, что дисфункция STN увеличивает импульсивность у людей, которым предъявлены два одинаково полезных стимула.[22]
Исследования показали, что субталамус - это экстрапирамидный центр. Он контролирует мышечные реакции, и повреждение может привести к гемибаллизму (сильному раскачиванию руки и ноги с одной стороны тела).[23]
Дополнительные изображения

Венечный отдел головного мозга непосредственно перед мостом. Субталамическое ядро обозначено как «Ядро Луйса».

Смотрите также
Рекомендации
- ^ Луйс, Жюль Бернар (1865). Recherches sur le système cérébro-spinal, ses fonctions, ses fonctions et ses maladies (На французском). Париж: Байер.
- ^ Афшарпур, С. (1985). «Световой микроскопический анализ субталамических нейронов крыс, пропитанных методом Гольджи». Журнал сравнительной неврологии. 236 (1): 1–13. Дои:10.1002 / cne.902360102. PMID 4056088.
- ^ Rafols, J. A .; Фокс, К. А. (1976). «Нейроны в субталамическом ядре приматов: исследование Гольджи и электронно-микроскопическое исследование». Журнал сравнительной неврологии. 168 (1): 75–111. Дои:10.1002 / cne.901680105. PMID 819471.
- ^ Ельник, Дж .; Першерон, Г. (1979). «Субталамические нейроны приматов: количественная и сравнительная анатомия». Неврология. 4 (11): 1717–1743. Дои:10.1016/0306-4522(79)90030-7. PMID 117397.
- ^ Levesque J.C .; Родитель А. (2005). «ГАМКергические интернейроны в субталамическом ядре человека». Двигательные расстройства. 20 (5): 574–584. Дои:10.1002 / mds.20374. PMID 15645534.
- ^ Кантерас Н.С., Шамма-Лагнадо С.Дж., Силва Б.А., Рикардо Дж.А. (апрель 1990 г.). «Афферентные связи субталамического ядра: комбинированное ретроградное и антероградное исследование пероксидазы хрена на крысах». Мозг Res. 513 (1): 43–59. Дои:10.1016 / 0006-8993 (90) 91087-В. PMID 2350684.
- ^ Cragg S.J .; Baufreton J .; Xue Y .; Болам Дж. П. и Беван М. Д. (2004). «Синаптическое высвобождение дофамина в субталамическом ядре». Европейский журнал нейробиологии. 20 (7): 1788–1802. Дои:10.1111 / j.1460-9568.2004.03629.x. PMID 15380000.
- ^ а б Наута Х.Дж., Коул М. (июль 1978 г.). «Эфферентные проекции субталамического ядра: авторадиографическое исследование на обезьянах и кошках». J. Comp. Neurol. 180 (1): 1–16. Дои:10.1002 / cne.901800102. PMID 418083.
- ^ а б Smith, Y .; Хазрати, L-N .; Родитель, А. (1990). «Эфферентные проекции субталамического ядра у беличьей обезьяны, как изучено методом антероградного отслеживания PHA-L». Журнал сравнительной неврологии. 294 (2): 306–323. Дои:10.1002 / cne.902940213. PMID 2332533.
- ^ Kita, H .; Чанг, Х. И Китай, С. (1983). «Морфология внутриклеточно меченых субталамических нейронов крысы: световой микроскопический анализ». Неврология. 215 (3): 245–257. Дои:10.1002 / cne.902150302. PMID 6304154.
- ^ Surmeier D.J.; Мерсер Дж. Н. И Чан С.С. (2005). «Автономные кардиостимуляторы в базальных ганглиях: кому вообще нужны возбуждающие синапсы?». Текущее мнение в нейробиологии. 15 (3): 312–318. Дои:10.1016 / j.conb.2005.05.007. PMID 15916893.
- ^ Levy R .; Hutchison W.D .; Лозано А.М. И Достровский Ю. (2000). «Высокочастотная синхронизация нейрональной активности в субталамическом ядре у пациентов с паркинсонизмом и тремором конечностей». Журнал неврологии. 20 (20): 7766–7775. Дои:10.1523 / JNEUROSCI.20-20-07766.2000. ЧВК 6772896. PMID 11027240.
- ^ Линтас А .; Силкис И.Г .; Albéri L .; Вилла A.E.P. (2012). «Дефицит дофамина увеличивает синхронизированную активность в субталамическом ядре крысы» (PDF). Исследование мозга. 1434 (3): 142–151. Дои:10.1016 / j.brainres.2011.09.005. PMID 21959175.
- ^ Пленц Д. и Китаи С. (1999). «Пейсмейкер базальных ганглиев, образованный субталамическим ядром и внешним бледным шаром». Природа. 400 (6745): 677–682. Дои:10.1038/23281. PMID 10458164.
- ^ Ельник, Дж. (2002). «Функциональная анатомия базальных ганглиев». Двигательные расстройства. 17 (Приложение 3): S15 – S21. Дои:10.1002 / mds.10138. PMID 11948751.
- ^ Sato, F .; Lavallée, P .; Левеск М. и Родитель А. (2000). "Одноаксонное отслеживание нейронов внешнего сегмента бледного шара у приматов". Журнал сравнительной неврологии. 417 (1): 17–31. Дои:10.1002 / (SICI) 1096-9861 (20000131) 417: 1 <17 :: AID-CNE2> 3.0.CO; 2-I. PMID 10660885.
- ^ Smith, Y .; Вичманн, Т. и ДеЛонг, М.Р. (1994). «Синаптическая иннервация нейронов во внутреннем паллидном сегменте субталамическим ядром и внешним паллидумом у обезьян». Журнал сравнительной неврологии. 343 (2): 297–318. Дои:10.1002 / cne.903430209. PMID 8027445.
- ^ Bevan M.D .; Мэджилл П.Дж .; Терман Д .; Болам Дж. П. и Уилсон С. Дж. (2002). «Двигайтесь в ритм: колебания в сети субталамического ядра - внешнего бледного шара». Тенденции в неврологии. 25 (10): 525–531. Дои:10.1016 / S0166-2236 (02) 02235-X. PMID 12220881.
- ^ Бекар Л .; Либионка В .; Tian G .; Xu Q .; Торрес А .; Ван Х .; Lovatt D .; Williams E .; Такано Т .; Schnermann J .; Бакос Р .; Недергаард М. (2008). «Аденозин имеет решающее значение для ослабления тремора, опосредованного глубокой стимуляцией мозга». Природа Медицина. 14 (1): 75–80. Дои:10,1038 / нм 1693. PMID 18157140.
- ^ Картер, Рита. Книга человеческого мозга. С. 58, 233.
- ^ L, молоток; Polosan M, Jaafari N, Baup N, Welter ML, Fontaine D; и другие. (2008). «Стимуляция субталамического ядра при тяжелом обсессивно-компульсивном расстройстве». Медицинский журнал Новой Англии. 359 (20): 2121–34. Дои:10.1056 / NEJMoa0708514. PMID 19005196.CS1 maint: несколько имен: список авторов (связь)
- ^ Франк, М .; Samanta, J .; Мустафа, А .; Шерман, С. (2007). «Держите лошадей: импульсивность, глубокая стимуляция мозга и лекарства при паркинсонизме». Наука. 318 (5854): 1309–12. Дои:10.1126 / science.1146157. PMID 17962524.
- ^ Брюс Х. Робинсон (2007). Биомедицина - Учебник для практиков акупунктуры и восточной медицины. Пресс для голубого мака. п. 126. ISBN 978-1-891845-38-3. LCCN 2006940894.
| Авторитетный контроль |
|---|