Кора головного мозга - Cerebral cortex
| Кора головного мозга | |
|---|---|
 Срез ткани мозга взрослой обезьяны макаки (Macaca mulatta). Кора головного мозга - это внешний слой, изображенный темно-фиолетовым цветом. Источник: BrainMaps.org | |
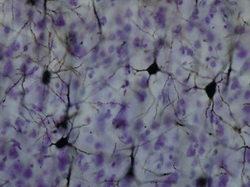 Окрашенный по Гольджи нейроны в коре | |
| подробности | |
| Часть | Головной мозг |
| Идентификаторы | |
| латинский | Кора головного мозга |
| MeSH | D002540 |
| NeuroNames | 39 |
| НейроЛекс МНЕ БЫ | birnlex_1494 |
| TA98 | A14.1.09.003 A14.1.09.301 |
| TA2 | 5527, 5528 |
| FMA | 61830 |
| Анатомические термины нейроанатомии | |
В кора головного мозга (множественное число кора), также известный как церебральная мантия,[1] это внешний слой нервная ткань из головной мозг из мозг в люди и другие млекопитающие. Кора головного мозга в основном состоит из шести слоев неокортекс, причем всего десять процентов состоит из аллокортекс.[2] Он разделен на два кора, посредством продольная трещина который разделяет головной мозг на левый и правый полушария головного мозга. Два полушария соединены корой головного мозга мозолистое тело. Кора головного мозга - самый большой участок нейронная интеграция в Центральная нервная система.[3] Он играет ключевую роль в внимание, восприятие, осведомленность, мысль, объем памяти, язык, и сознание.
У большинства млекопитающих, за исключением мелких млекопитающих с маленьким мозгом, кора головного мозга имеет складчатую форму, обеспечивая большую площадь поверхности в ограниченном объеме мозга. череп. Помимо минимизации объема мозга и черепа, корковая складка имеет решающее значение для проводка мозга и его функциональная организация. У млекопитающих с маленьким мозгом складчатость отсутствует, кора гладкая.[4][5]
Складка или гребень в коре называется извилина (множественные извилины) и канавка называется борозда (множественное число борозд). Эти поверхностные извилины появляются во время развитие плода и продолжают созревать после рождения в процессе гирификация. в человеческий мозг большая часть коры головного мозга не видна снаружи, а скрыта в бороздах,[6] и островковая кора полностью скрыт. Основные борозды и извилины отмечают подразделения головного мозга на доли мозга.
От 14 до 16 миллиардов нейроны в коре головного мозга. Они организованы в горизонтальные слои и радиально в корковые столбики и мини-столбцы. Корковые области выполняют определенные функции, такие как движение в моторная кора, и взгляд в зрительная кора.
Структура

Кора головного мозга представляет собой внешнее покрытие поверхностей полушарий головного мозга и складывается в вершины, называемые извилины, а канавки называются борозды. в человеческий мозг он имеет толщину от двух до трех или четырех миллиметров,[7] и составляет 40 процентов массы мозга.[3] 90 процентов коры головного мозга представляет собой шестислойную неокортекс а остальные 10 процентов составляют аллокортекс.[3] В коре от 14 до 16 миллиардов нейронов.[3] и они организованы радиально в корковые столбики, и мини-столбцы, в горизонтально организованных слоях коры.[8][9]
Около двух третей кортикальной поверхности находится в бороздах, а островковая кора полностью скрыт. Кора головного мозга наиболее толстая на вершине извилины и самая тонкая на нижней части борозды.[10]
Складки
Кора головного мозга сложена таким образом, что позволяет покрывать большую площадь поверхности. нервная ткань вписаться в рамки нейрокраниум. Когда раскрывается в человеке, каждый полушарный кора головного мозга имеет общую площадь поверхности около 0,12 квадратных метра (1,3 квадратных фута).[11] Складка направлена внутрь от поверхности мозга, а также присутствует на медиальной поверхности каждого полушария в пределах продольная трещина. У большинства млекопитающих кора головного мозга состоит из пиков, известных как извилины, и впадин или бороздок, известных как борозды. Некоторые мелкие млекопитающие, включая некоторых мелких грызуны иметь гладкую мозговую поверхность без гирификация.[5]
Доли
Более крупные борозды и извилины отмечают подразделения коры головного мозга на доли мозга.[7] Есть четыре основных доли: Лобная доля, теменная доля, височная доля, и затылочная доля. В островковая кора часто включается как островная доля.[12] В лимбическая доля это край коры на медиальной стороне каждого полушария, который также часто присутствует.[13] Также описаны три доли головного мозга: парацентральная долька, то верхняя теменная долька, а нижняя теменная долька.
Толщина
У видов млекопитающих более крупный мозг (в абсолютном выражении, а не только по отношению к размеру тела), как правило, имеет более толстую корку.[14] Самые маленькие млекопитающие, такие как землеройки имеют толщину неокортекса около 0,5 мм; те, у кого самый большой мозг, такие как люди и финвалы, имеют толщину 2–4 мм.[3][7] Существует примерно логарифмический взаимосвязь между массой мозга и толщиной коры.[14]Магнитно-резонансная томография головного мозга (МРТ) позволяет измерить толщину коры головного мозга человека и связать ее с другими показателями. Толщина различных областей коры различается, но в целом сенсорная кора тоньше моторной коры.[15] Одно исследование обнаружило некоторую положительную связь между толщиной коркового слоя и интеллект.[16]Другое исследование показало, что соматосенсорная кора толще в мигрень больных, хотя неизвестно, является ли это результатом приступов мигрени или их причиной.[17][18]В более позднем исследовании с участием большего количества пациентов сообщается об отсутствии изменений в толщине коркового слоя у страдающих мигренью.[19]Генетическое заболевание коры головного мозга, при котором уменьшение складчатости в определенных областях приводит к микрогирс, где четыре слоя вместо шести, в некоторых случаях считается связанным с дислексия.[20]
Слои неокортекса
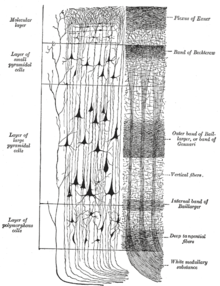
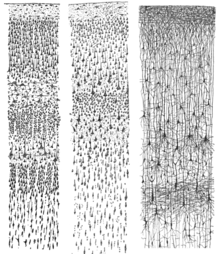

В неокортекс состоит из шести корковых слоев, пронумерованных от I до VI от самого дальнего (прилегающего к pia mater ) к самому внутреннему (примыкающему к белое вещество ). Каждый слой имеет характерное распределение разных нейронов и их связи с другими корковыми и подкорковыми областями. Есть прямые связи между различными областями коры и косвенные связи через таламус.
Один из ярких примеров коркового слоя это линия Дженнари в первичная зрительная кора. Это полоса более белой ткани, которую можно увидеть невооруженным глазом на дне известковая борозда затылочной доли. Линия Дженнари состоит из аксоны принося визуальную информацию из таламус в слой IV зрительная кора.
Окрашивание поперечные сечения коры головного мозга для выявления положения тел нейрональных клеток и интракортикальных трактов аксонов позволили нейроанатомам в начале 20-го века произвести подробное описание ламинарная структура коры у разных видов. Работа Корбинян Бродманн (1909) установили, что неокортекс млекопитающих (и большинство других областей коры) последовательно делится на шесть слоев.
Слой I
Слой I - это молекулярный слой, и содержит несколько разбросанных нейронов, в том числе ГАМКергический нейроны шиповника.[21] Слой I в основном состоит из продолжений апикального дендритный пучки пирамидные нейроны и горизонтально ориентированные аксоны, а также глиальные клетки.[22] Во время разработки Клетки Кахаля-Ретциуса[23] и клетки субпиального зернистого слоя[24] присутствуют в этом слое. Кроме того, некоторые колючие звездчатые клетки можно найти здесь. Считается, что входы в апикальные пучки имеют решающее значение для Обратная связь взаимодействия в коре головного мозга, участвующие в ассоциативном обучении и внимании.[25] Когда-то считалось, что вход в слой I исходит из самой коры,[26] теперь стало понятно, что слой I через мантию коры головного мозга получает существенный вклад от матрица или клетки таламуса М-типа[27] (в отличие от ядро или C-типа, которые переходят на уровень IV).[28]
Слой II
Слой II, внешний зернистый слой, содержит небольшие пирамидные нейроны и многочисленные звездчатые нейроны.
Слой III
Слой III, внешний пирамидальный слой, содержит преимущественно мелкие и средние пирамидные нейроны, а также непирамидные нейроны с вертикально ориентированными внутрикортикальными аксонами; слои с I по III являются основной мишенью межполушарных кортикортикальных афференты, а слой III является основным источником корково-коркового эфференты.
Слой IV
Слой IV, внутренний зернистый слой, содержит разные типы звездчатый и пирамидных клеток, и является основной мишенью таламокортикальные афференты из нейронов таламуса типа С (стержневого типа)[28] а также внутриполушарные кортикокортикальные афференты. Слои над слоем IV также называются супрагранулярными слоями (слои I-III), тогда как слои ниже называются инфрагранулярными слоями (слои V и VI).
Слой V
Слой V, внутренний пирамидальный слой, содержит большие пирамидные нейроны. Аксоны из них покидают кору и соединяются с подкорковыми структурами, включая базальный ганглий. В первичной моторной коре лобной доли слой V содержит гигантские пирамидальные клетки, называемые Клетки Беца, аксоны которого проходят через внутренняя капсула, то мозговой ствол, и спинной мозг, образующий кортикоспинальный тракт, который является основным путем произвольного моторного контроля.
Слой VI
Слой VI, полиморфный или многоформный слой, содержит несколько больших пирамидных нейронов и множество маленьких веретенообразных пирамидных и многоформных нейронов; слой VI посылает эфферентные волокна в таламус, устанавливая очень точное взаимное соединение между корой и таламусом.[29] То есть нейроны слоя VI из одного коркового столбца соединяются с нейронами таламуса, которые обеспечивают вход в тот же кортикальный столбец. Эти связи являются одновременно возбуждающими и тормозящими. Нейроны посылают возбуждающий волокна к нейронам в таламусе, а также отправляют коллатерали в ретикулярное ядро таламуса это подавлять эти самые нейроны таламуса или соседние с ними.[30] Одна из теорий состоит в том, что, поскольку тормозящий выход снижается на холинергический вход в кору головного мозга, это обеспечивает мозговой ствол с регулируемым "регулятором усиления для реле лемнисковый входы ».[30]
Столбцы
Корковые слои не просто накладываются друг на друга; существуют характерные связи между различными слоями и типами нейронов, охватывающими всю толщину коры. Эти корковые микросхемы сгруппированы в корковые столбики и мини-столбцы.[31] Было высказано предположение, что миниколонки являются основными функциональными единицами коры.[32] В 1957 г. Вернон Маунткасл показали, что функциональные свойства коры резко меняются между латерально соседними точками; однако они непрерывны в направлении, перпендикулярном поверхности. Более поздние работы предоставили доказательства наличия функционально различных кортикальных столбов в зрительной коре (Hubel и Визель, 1959),[33] слуховая кора и ассоциативная кора.
Области коры, в которых отсутствует слой IV, называются зернистый. Области коры, которые имеют только рудиментарный слой IV, называются дисгранулярными.[34] Обработка информации в каждом слое определяется разной временной динамикой по сравнению с таковой в слоях II / III, имеющих медленное 2Гц колебание а в слое V - быстрые колебания 10–15 Гц.[35]
Типы коры
Исходя из различий в ламинарная организация кору головного мозга можно разделить на два типа: большая площадь неокортекс который имеет шесть слоев ячеек и гораздо меньшую площадь аллокортекс который имеет три или четыре слоя:[2]
- Неокортекс также известен как изокортекс или неопаллий и является частью зрелой коры головного мозга с шестью отдельными слоями. Примеры областей неокортекса включают зернистую первичная моторная кора, а полосатая первичная зрительная кора. Неокортекс имеет два подтипа: истинная изокортекс и произокортекс который является переходной областью между изокортексом и областями периаллокортекса.
- Аллокортекс - это часть коры головного мозга с тремя или четырьмя слоями и имеет три подтипа: палеокортекс с тремя корковыми пластинками, архикортекс который имеет четыре или пять, а также переходную область, примыкающую к аллокортексу, периаллокортекс. Примерами allocortex являются обонятельная кора и гиппокамп.
Между неокортексом и аллокортексом существует переходная область, называемая паралимбическая кора, где слои 2, 3 и 4 объединены. Эта область включает произокортекс неокортекса и периаллокортекс аллокортекса. Кроме того, кору головного мозга можно разделить на четыре доли: the Лобная доля, височная доля, то теменная доля, а затылочная доля, названные от их вышележащих костей черепа.
Кровоснабжение и дренаж

Кровоснабжение коры головного мозга является частью мозговое кровообращение. Церебральные артерии поставлять кровь, которая перфузии головной мозг. Эта артериальная кровь переносит кислород, глюкозу и другие питательные вещества в кору. Церебральные вены слить дезоксигенированную кровь и метаболические отходы, включая углекислый газ, обратно в сердце.
Основными артериями, снабжающими кору головного мозга, являются передняя мозговая артерия, то средняя мозговая артерия, а задняя мозговая артерия. Передняя мозговая артерия снабжает передние отделы головного мозга, включая большую часть лобной доли. Средняя мозговая артерия снабжает кровью теменные доли, височные доли и части затылочных долей. Средняя мозговая артерия разделяется на две ветви, чтобы снабжать левое и правое полушария, где они разветвляются дальше. Задняя мозговая артерия снабжает затылочные доли.
В круг Уиллиса является основной кровеносной системой, которая отвечает за кровоснабжение головного мозга и коры головного мозга.

Развитие
В пренатальное развитие коры головного мозга - сложный и точно настроенный процесс, называемый кортикогенез под влиянием взаимодействия генов и окружающей среды.[36]
Нервная трубка
Кора головного мозга развивается из самой передней части, области переднего мозга, нервная трубка.[37][38] В нервная пластинка складывается и закрывается, чтобы сформировать нервная трубка. Из полости внутри нервной трубки развивается желудочковая система, а из нейроэпителиальные клетки его стен, нейроны и глия нервной системы. Самая передняя (передняя или черепная) часть нервной пластинки, передний мозг, что очевидно перед нейруляция начинается, дает начало полушариям головного мозга, а затем и коре головного мозга.[39]
Развитие корковых нейронов
Корковые нейроны генерируются внутри желудочковая зона, сразу после желудочки. Сначала эта зона содержит нервные стволовые клетки, этот переход к радиальные глиальные клетки –Клетки-предшественники, которые делятся с образованием глиальных клеток и нейронов.[40]
Радиальная глия
Кора головного мозга состоит из гетерогенной популяции клеток, которые дают начало различным типам клеток. Большинство этих клеток происходит от радиальная глия миграции, которые формируют различные типы клеток неокортекса, и это период, связанный с увеличением нейрогенез. Точно так же процесс нейрогенеза регулирует ламинирование с образованием различных слоев коры. Во время этого процесса происходит усиление ограничения клеточной судьбы, которое начинается с более раннего прародители давая начало любому типу клеток в коре головного мозга и более поздним предшественникам, давая начало только нейроны поверхностных слоев. Эта дифференцированная судьба клеток создает в коре головного мозга вывернутую топографию с более молодыми нейронами в поверхностных слоях и старыми нейронами в более глубоких слоях. Кроме того, ламинарные нейроны останавливаются в S или Фаза G2 чтобы дать четкое различие между разными корковыми слоями. Ламинарная дифференцировка не полностью завершается до момента рождения, поскольку во время развития ламинарные нейроны все еще чувствительны к внешним сигналам и сигналам окружающей среды.[41]
Хотя большинство клеток, составляющих кору, происходит локально из радиальной глии, существует подмножество популяции нейронов, которые мигрировать из других регионов. Радиальная глия дает начало нейронам пирамидальной формы и используется глутамат как нейротрансмиттер, однако эти мигрирующие клетки вносят свой вклад в нейроны звездчатой формы, которые используют ГАМК как их главный нейротрансмиттер. Эти ГАМКергические нейроны генерируются клетками-предшественниками в возвышение медиального ганглия (MGE), которые перемещаются по касательной к коре через субвентрикулярная зона. Эта миграция ГАМКергических нейронов особенно важна, поскольку Рецепторы ГАМК возбуждают в процессе развития. Это возбуждение в первую очередь вызвано потоком ионов хлорида через рецептор ГАМК, однако у взрослых концентрация хлоридов смещается, вызывая поступление внутрь хлорида, которое гиперполяризует постсинаптические нейроны.[42]Глиальные волокна, образующиеся в первых отделах клеток-предшественников, ориентированы радиально, охватывая толщину коры от желудочковая зона к внешнему, пиал поверхность, и обеспечивают основу для миграции нейронов наружу от желудочковая зона.[43][44]
При рождении очень мало дендриты присутствует на теле клетки коркового нейрона, а аксон не развит. В течение первого года жизни количество дендритов резко увеличивается, так что они могут вместить до ста тысяч человек. синаптические связи с другими нейронами. Аксон может развиваться и простираться далеко от тела клетки.[45]
Асимметричное деление
Первые деления клеток-предшественников симметричны, что в каждом случае дублирует общее количество клеток-предшественников. митотический цикл. Затем некоторые клетки-предшественники начинают асимметрично делиться, производя одну постмитотическую клетку, которая мигрирует по радиальным глиальным волокнам, оставляя желудочковая зона, и одна клетка-предшественник, которая продолжает делиться до конца развития, когда она дифференцируется в глиальная клетка или эпендимная клетка. Поскольку Фаза G1 из митоз является удлиненным, что рассматривается как избирательное удлинение клеточного цикла, новорожденные нейроны мигрируют в более поверхностные слои коры.[46] Мигрирующие дочерние клетки становятся пирамидные клетки коры головного мозга.[47] Процесс развития упорядочен во времени и регулируется сотнями генов и эпигенетические регуляторные механизмы.[48]
Слой организации

В слоистая структура зрелой коры головного мозга формируется в процессе развития. Первые сформированные пирамидные нейроны мигрируют из желудочковая зона и субвентрикулярная зона, вместе с катушка -производство Нейроны Кахаля – Ретциуса, от предварительная пластина. Затем когорта нейронов, мигрирующих в середину препластинки, делит этот переходный слой на поверхностный. краевая зона, который станет слоем I зрелого неокортекса, и плита,[49] формируя средний слой, называемый корковая пластинка. Эти клетки образуют глубокие слои зрелой коры, пятый и шестой слои. Позже рожденные нейроны мигрируют радиально в кортикальную пластинку, минуя нейроны глубокого слоя, и становятся верхними слоями (двумя-четырьмя). Таким образом, слои коры создаются в порядке «наизнанку».[50] Единственное исключение из этой вывернутой наизнанку последовательности нейрогенез находится в слое I приматы, в котором, в отличие от грызуны, нейрогенез продолжается на протяжении всего периода кортикогенез.[51]
Кортикальный паттерн
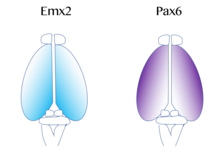
Карта функциональных областей коры, которые включают первичную моторную и зрительную кору, происходит отПротокарта ',[52] который регулируется молекулярными сигналами, такими как фактор роста фибробластов FGF8 в начале эмбрионального развития.[53][54] Эти сигналы регулируют размер, форму и положение корковых областей на поверхности коркового зачатка, частично за счет регулирования градиентов фактор транскрипции выражение через процесс, называемый кортикальный паттерн. Примеры таких факторов транскрипции включают гены EMX2 и PAX6.[55] Вместе оба факторы транскрипции образуют противоположный градиент выражения. Pax6 высоко выражено на ростральный боковой полюс, а Emx2 сильно выражено в каудомедиальный столб. Установление этого градиента важно для правильного развития. Например, мутации в Pax6 может вызвать расширение уровней экспрессии Emx2 за пределы его нормального домена экспрессии, что в конечном итоге приведет к расширению областей, обычно происходящих из каудальной медиальной коры, таких как зрительная кора. Напротив, если происходят мутации в Emx2, это может вызвать расширение экспрессирующего Pax6 домена и привести к лобной и моторный кортикальный расширение регионов. Поэтому исследователи считают, что подобные градиенты и центры сигнализации рядом с корой может способствовать региональной экспрессии этих факторов транскрипции.[42]Два хорошо изученных паттернирующих сигнала для коры включают: FGF и ретиноевая кислота. Если FGF неправильно выраженный в разных областях развивающейся коры, кортикальный паттерн нарушается. В частности, когда Fgf8 увеличивается в передний полюс, Emx2 подавленный и каудальный происходит сдвиг в корковой области. В конечном итоге это вызывает расширение ростральных областей. Следовательно, Fgf8 и др. FGFs играют роль в регуляции экспрессии Emx2 и Pax6 и представляют, как кора головного мозга может становиться специализированной для различных функций.[42]
Быстрое расширение площади поверхности коры регулируется количеством самообновления радиальные глиальные клетки и частично регулируется FGF и Notch гены.[56] В период коркового нейрогенеза и формирования слоев у многих высших млекопитающих начинается процесс гирификация, образующий характерные складки коры головного мозга.[57][58] Гирификация регулируется ДНК-связанным белком Trnp1[59] и FGF и SHH сигнализация[60][61]
Эволюция
Из всех областей мозга кора головного мозга демонстрирует наибольшие эволюционные вариации и развивалась совсем недавно.[5] В отличие от очень консервативной схемы продолговатый мозг Например, который выполняет важные функции, такие как регулирование частоты сердечных сокращений и дыхания, многие области коры головного мозга не являются строго необходимыми для выживания. Таким образом, эволюция коры головного мозга привела к появлению и модификации новых функциональных областей - особенно ассоциативных областей, которые не получают непосредственного ввода извне коры.[5]
Ключевая теория корковой эволюции воплощена в гипотеза радиального устройства и связанные Протокарта гипотеза, впервые предложенная Ракичем.[62] Эта теория утверждает, что новые области коры формируются путем добавления новых радиальных единиц, что достигается в стволовая клетка уровень. Гипотеза протокарты утверждает, что клеточная и молекулярная идентичность и характеристики нейронов в каждой области коры определяются корковой стволовые клетки, известный как радиальные глиальные клетки, в изначальной карте. Эта карта контролируется секретной сигнализацией. белки и вниз по течению факторы транскрипции.[63][64][65]
Функция
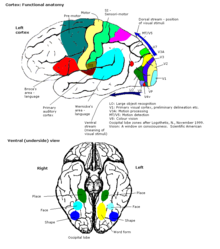
Подключения
Кора головного мозга связана с различными подкорковыми структурами, такими как таламус и базальный ганглий отправляя им информацию вместе эфферентные связи и получать от них информацию через афферентные связи. Большая часть сенсорной информации направляется в кору головного мозга через таламус. Обонятельная информация, однако, проходит через обонятельная луковица в обонятельную кору (грушевидная кора ). Большинство соединений идет от одной области коры к другой, а не от подкорковых областей; Брайтенберг и Schüz (1998) утверждают, что в первичных сенсорных областях, на кортикальном уровне, где заканчиваются входные волокна, до 20% синапсов снабжается экстракортикальными афферентами, но что в других областях и других слоях этот процент, вероятно, будет намного ниже. .[66]
Корковые области


Вся кора головного мозга была разделена на 52 различные области на ранних этапах исследования. Корбинян Бродманн. Эти области, известные как Площади Бродмана, основаны на их цитоархитектура но также относятся к различным функциям. Примером может служить зона Бродмана 17, которая является первичная зрительная кора.
В более общем плане кора обычно описывается как состоящая из трех частей: сенсорной, моторной и ассоциативной областей.
Сенсорные зоны
Сенсорные области - это области коры, которые получают и обрабатывают информацию от чувства. Части коры, которые получают сенсорные сигналы от таламус называются первичными сенсорными зонами. Чувства зрения, слуха и осязания обслуживаются первичной зрительной корой, первичной слуховая кора и первичная соматосенсорная кора соответственно. Как правило, два полушария получают информацию с противоположной (контралатеральной) стороны тела. тело. Например, правая первичная соматосенсорная кора получает информацию от левых конечностей, а правая зрительная кора получает информацию от левого зрительного нерва. поле. Организация сенсорных карт в коре головного мозга отражает организацию соответствующего сенсорного органа, известного как топографическая карта. Соседние точки в праймериз зрительная кора, например, соответствуют соседним точкам в сетчатка. Эта топографическая карта называется ретинотопная карта. Таким же образом существует тонотопическая карта в первичной слуховой коре и соматотопическая карта в первичной сенсорной коре. Эта последняя топографическая карта тела на задняя центральная извилина был проиллюстрирован как деформированное человеческое представление, соматосенсорная гомункул, где размер различных частей тела отражает относительную плотность их иннервации. Области с большой сенсорной иннервацией, такие как кончики пальцев и губы, требуют большей площади коры для обработки более тонких ощущений.
Двигательные области
Двигательные области расположены в обоих полушариях коры. Двигательные области очень тесно связаны с контролем произвольных движений, особенно мелких фрагментированных движений, выполняемых рукой. Правая половина моторной области контролирует левую сторону тела, и наоборот.
Две области коры обычно называют моторными:
- Первичная моторная кора, который выполняет произвольные движения[нужна цитата ]
- Дополнительные моторные области и премоторная кора, который Выбрать произвольные движения.[нужна цитата ]
Кроме того, моторные функции описаны для:
- Задняя теменная кора, который направляет произвольные движения в пространстве
- Дорсолатеральная префронтальная кора, который решает, какие произвольные движения выполнять в соответствии с инструкциями, правилами и самопроизвольными мыслями более высокого порядка.
Прямо под корой головного мозга находятся взаимосвязанные подкорковые массы серого вещества, называемые базальный ганглий (или ядра). Базальные ганглии получают сигнал от черной субстанции среднего мозга и моторных областей коры головного мозга и посылают сигналы обратно в оба этих места. Они участвуют в двигательном контроле. Они находятся сбоку от таламуса. Основными компонентами базальных ганглиев являются хвостатое ядро, то скорлупа, то бледный шар, то черная субстанция, то прилежащее ядро, а субталамическое ядро. Скорлупа и бледный шар также известны под общим названием чечевицеобразное ядро, потому что вместе они образуют линзовидное тело. Скорлупа и хвостатое ядро также собирательно называют полосатое тело после их полосатого вида.[67][68]
Области ассоциации

Области ассоциации - это части коры головного мозга, не принадлежащие к первичным областям. Они функционируют, чтобы произвести значимое перцептивный опыт мира, позволяют нам эффективно взаимодействовать и поддерживают абстрактное мышление и язык. В теменный, временный, и затылочные доли - все они расположены в задней части коры головного мозга - объединяют сенсорную информацию и информацию, хранящуюся в памяти. В Лобная доля Комплекс префронтальных ассоциаций участвует в планировании действий и движений, а также в абстрактном мышлении. В глобальном масштабе зоны ассоциации организованы как распределенные сети.[69] Каждая сеть соединяет области, распределенные по широко разнесенным областям коры. Отдельные сети расположены рядом друг с другом, образуя сложную серию переплетенных сетей. Конкретная организация ассоциативных сетей обсуждается с доказательствами взаимодействий, иерархических отношений и конкуренции между сетями.[70]
У людей ассоциативные сети особенно важны для языковой функции. В прошлом предполагалось, что языковые способности локализованы в Площадь Брока в областях левых нижняя лобная извилина, BA44 и BA45, для языкового выражения и в Площадь Вернике BA22, для языкового приема. Однако было показано, что процессы языкового выражения и восприятия происходят не только в структурах вокруг боковая борозда, включая лобную долю, базальный ганглий, мозжечок, и мосты.[71]
Клиническое значение
Нейродегенеративные заболевания, такие как Болезнь Альцгеймера и Болезнь Лафора, показать в качестве маркера атрофию серого вещества коры головного мозга.[73]
Другой заболевания центральной нервной системы включают неврологические расстройства такие как эпилепсия, двигательные расстройства, и трудности в речи (афазия ).
Повреждение мозга от болезни или травмы, может включать повреждение определенной доли, например, заболевание лобной доли, и связанные функции будут затронуты. В гематоэнцефалический барьер который служит для защиты мозга от инфекции, может быть нарушен, позволяя проникнуть в патогены.
В развивающийся плод чувствителен к ряду факторов окружающей среды, которые могут вызвать врожденные дефекты и проблемы в дальнейшем развитии. Например, употребление алкоголя матерью может вызвать расстройство алкогольного спектра плода.[74] Другими факторами, которые могут вызвать нарушения нервного развития, являются: токсиканты такие как наркотики, и воздействие радиация как из Рентгеновские лучи. Инфекции также могут влиять на развитие коры головного мозга. Вирусная инфекция - одна из причин лиссэнцефалия, что приводит к гладкой коре без гирификация.
Тип электрокортикография называется картирование корковой стимуляции это инвазивная процедура, которая включает размещение электроды непосредственно на обнаженный мозг, чтобы локализовать функции определенных областей коры. Он используется в клинических и терапевтических целях, включая предоперационное картирование.[75]
Гены, связанные с корковыми нарушениями
Существует ряд генетических мутаций, которые могут вызывать широкий спектр генетические нарушения коры головного мозга, в том числе микроцефалия, шизэнцефалия и виды лиссэнцефалия.[76] Хромосомные аномалии может также вызвать ряд расстройства нервного развития такие как синдром ломкой Х-хромосомы и Синдром Ретта.
MCPH1 коды для микроцефалин, и нарушения в этом и в ASPM связаны с микроцефалией.[76] Мутации в гене NBS1 это коды для нибрин может вызвать Синдром перелома Неймегена, характеризующийся микроцефалией.[76]
Мутации в EMX2,[77] и COL4A1 связаны с шизэнцефалия,[78] состояние, при котором отсутствуют большие части полушарий головного мозга.
История
В 1909 г. Корбинян Бродманн выделили различные области неокортекса на основе цитоархитектурных различий и разделили кору головного мозга на 52 области.[79]
Рафаэль Лоренте де Но, студент Сантьяго Рамон-и-Кахаль идентифицировали более 40 различных типов корковых нейронов на основе распределения их дендритов и аксонов.[79]
Другие животные
Кора головного мозга происходит от паллий, слоистая структура, обнаруженная в передний мозг из всех позвоночные. Основная форма паллия - это цилиндрический слой, охватывающий желудочки, заполненные жидкостью. По окружности цилиндра расположены четыре зоны: дорсальный паллий, медиальный паллий, вентральный паллий и латеральный паллий, которые, как считается, соответственно образуют неокортекс, гиппокамп, миндалина, и обонятельная кора.
До недавнего времени аналог коры головного мозга у беспозвоночных не обнаруживался. Однако исследование, опубликованное в журнале Ячейка в 2010 году на основе профилей экспрессии генов сообщалось о сильном сродстве между корой головного мозга и грибовидные тела из тряпка Platynereis dumerilii.[80] Грибные тела - это структуры в мозге многих типов червей и членистоногих, которые, как известно, играют важную роль в обучении и памяти; генетические данные указывают на общее эволюционное происхождение и, следовательно, указывают на то, что происхождение самых ранних предшественников коры головного мозга восходит к ранним Докембрийский эпоха.
Дополнительные изображения

Моторные и сенсорные области коры головного мозга

Моторные и сенсорные области коры головного мозга


Смотрите также
использованная литература
- ^ "церебральная мантия". TheFreeDictionary.com.
- ^ а б Строминджер, Норман Л .; Демарест, Роберт Дж .; Лемле, Лоис Б. (2012). "Кора головного мозга". Нервная система человека Нобака, седьмое издание. Humana Press. С. 429–451. Дои:10.1007/978-1-61779-779-8_25. ISBN 978-1-61779-778-1.
- ^ а б c d е Саладин, Кеннет (2011). Человеческая анатомия (3-е изд.). Макгроу-Хилл. С. 416–422. ISBN 9780071222075.
- ^ Фернандес, V; Ллинарес-Бенадеро, C; Боррелл, В. (17 мая 2016 г.). «Расширение и складывание коры головного мозга: чему мы научились?». Журнал EMBO. 35 (10): 1021–44. Дои:10.15252 / embj.201593701. ЧВК 4868950. PMID 27056680.
- ^ а б c d Ракич, П. (октябрь 2009 г.). «Эволюция неокортекса: взгляд из биологии развития». Обзоры природы Неврология. 10 (10): 724–35. Дои:10.1038 / nrn2719. ЧВК 2913577. PMID 19763105.
- ^ Принципы нейронауки (4-е изд.). McGraw-Hill, Отдел медицинских профессий. 2000-01-05. ISBN 978-0838577011.
- ^ а б c Робертс, П. (1992). Нейроанатомия (3-е изд.). Springer-Verlag. С. 86–92. ISBN 9780387977775.
- ^ Лодато, Симона; Арлотта, Паола (13 ноября 2015 г.). «Создание нейронального разнообразия в коре головного мозга млекопитающих». Ежегодный обзор клеточной биологии и биологии развития. 31 (1): 699–720. Дои:10.1146 / annurev-cellbio-100814-125353. ЧВК 4778709. PMID 26359774.
Функциональные столбцы были впервые определены в коре Маунткаслом (Mountcastle, 1957), который предложил столбчатую гипотезу, согласно которой кора состоит из дискретных модульных столбцов нейронов, характеризующихся последовательным профилем связности.
- ^ Ансен-Уилсон, LJ; Липинский, Р. Дж. (Январь 2017 г.). «Взаимодействие генов и окружающей среды в развитии и дисфункции корковых интернейронов: обзор доклинических исследований». Нейротоксикология. 58: 120–129. Дои:10.1016 / j.neuro.2016.12.002. ЧВК 5328258. PMID 27932026.
- ^ Карпентер (1985). Основной текст нейроанатомии (3-е изд.). Уильямс и Уилкинс. С. 348–358. ISBN 978-0683014556.
- ^ Торо, Роберто; Перрон, Мишель; Пайк, Брюс; Ричер, Луи; Вейлетт, Сюзанна; Паусова, Зденка; Паус, Томаш (01.10.2008). «Размер мозга и складка коры головного мозга человека». Кора головного мозга. 18 (10): 2352–2357. Дои:10.1093 / cercor / bhm261. ISSN 1047-3211. PMID 18267953.
- ^ Nieuwenhuys, R (2012). «Островная кора: обзор». Прогресс в исследованиях мозга. 195: 123–63. Дои:10.1016 / B978-0-444-53860-4.00007-6. PMID 22230626.
- ^ Tortora, G; Дерриксон, Б. (2011). Основы анатомии и физиологии (13-е изд.). Вайли. п. 549. ISBN 9780470646083.
- ^ а б Nieuwenhuys R, Donkelaar HJ, Nicholson C (1998). Центральная нервная система позвоночных, Том 1. Springer. С. 2011–2012. ISBN 978-3-540-56013-5.
- ^ Фритьоф Круггель; Мартина К. Брюкнер; Томас Арендт; Кристофер Дж. Виггинс; Д. Ив фон Крамон (2003). «Анализ тонкой структуры неокортекса». Анализ медицинских изображений. 7 (3): 251–264. Дои:10.1016 / S1361-8415 (03) 00006-9. HDL:11858 / 00-001M-0000-0010-9C60-3. PMID 12946467.
- ^ Кэтрин Л. Нарр; Роджер П. Вудс; Пол М. Томпсон; Филип Шешко; Дилберт Робинсон; Теодора Димчева; Мала Гурбани; Артур В. Тога; Роберт М. Билдер (2007). «Связь между IQ и региональной толщиной серого вещества коры у здоровых взрослых». Кора головного мозга. 17 (9): 2163–2171. Дои:10.1093 / cercor / bhl125. PMID 17118969.
- ^ Александр Ф.М. DaSilva; Кристина Гранзьера; Джош Снайдер; Нучин Хаджихани (2007). «Утолщение соматосенсорной коры у больных мигренью». Неврология. 69 (21): 1990–1995. Дои:10.1212 / 01.wnl.0000291618.32247.2d. ЧВК 3757544. PMID 18025393.
- ^ Кэтрин Пэддок (2007-11-20). «У больных мигренью более толстая кора головного мозга». Медицинские новости сегодня. В архиве из оригинала от 11.05.2008.
- ^ Датте Р., Детре Дж. А. и др. (Октябрь 2011 г.). «Отсутствие изменений толщины коркового слоя у больных мигренью». Цефалгия. 31 (14): 1452–8. Дои:10.1177/0333102411421025. ЧВК 3512201. PMID 21911412.
- ^ Хабиб М (2000). «Неврологические основы дислексии развития: обзор и рабочая гипотеза». Мозг. 123 (12): 2373–99. Дои:10.1093 / мозг / 123.12.2373. PMID 11099442.
- ^ «Ученые определили новый вид клетки человеческого мозга». Институт Аллена. 27 августа 2018.
- ^ Шипп, Стюарт (17.06.2007). «Строение и функции коры головного мозга». Текущая биология. 17 (12): R443–9. Дои:10.1016 / j.cub.2007.03.044. ЧВК 1870400. PMID 17580069.
- ^ Мейер, Гундела; Goffinet, André M .; Фэйрен, Альфонсо (1999). «Особая статья: Что такое клетка Кахаля – Ретциуса? Переоценка классического типа клеток на основе недавних наблюдений в развивающемся неокортексе». Кора головного мозга. 9 (8): 765–775. Дои:10.1093 / cercor / 9.8.765. PMID 10600995. В архиве из оригинала от 21.02.2015.
- ^ Иудаш, Милош; Плетикос, Миховил (2010). «Открытие субпиального гранулярного слоя в коре головного мозга человека». Трансляционная нейронаука. 1 (3): 255–260. Дои:10.2478 / v10134-010-0037-4. S2CID 143409890.
- ^ Гилберт CD, Сигман М (2007). «Состояния мозга: влияние сверху вниз на сенсорную обработку». Нейрон. 54 (5): 677–96. Дои:10.1016 / j.neuron.2007.05.019. PMID 17553419.
- ^ Коллер Л. (1995). «Слой I первичной сенсорной неокортекса: где сходится нисходящий сверху вниз». Behav Brain Res. 71 (1–2): 163–70. Дои:10.1016/0166-4328(95)00032-1. PMID 8747184. S2CID 4015532.
- ^ Рубио-Гарридо П., Перес-де-Манцо Ф., Порреро С., Галазо М.Дж., Класка Ф (2009). «Вход таламуса в дистальные апикальные дендриты в неокортикальном слое 1 массивный и сильно конвергентный». Кора головного мозга. 19 (10): 2380–95. Дои:10.1093 / cercor / bhn259. PMID 19188274.
- ^ а б Джонс EG (1998). «Точка зрения: ядро и матрица таламической организации». Неврология. 85 (2): 331–45. Дои:10.1016 / S0306-4522 (97) 00581-2. PMID 9622234. S2CID 17846130.
- ^ Кройцфельдт, О. 1995. Cortex Cerebri. Springer-Verlag.
- ^ а б Лам Ю.В., Шерман С.М. (2010). «Функциональная организация соматосенсорного коркового слоя 6, обратная связь с таламусом». Кора головного мозга. 20 (1): 13–24. Дои:10.1093 / cercor / bhp077. ЧВК 2792186. PMID 19447861.
- ^ Сузуки, И.К .; Хирата, Т. (январь 2013 г.). «Неокортикальный нейрогенез - это не совсем« нео »: новая эволюционная модель, полученная на основе сравнительного исследования паллиального развития цыплят» (PDF). Развитие, рост и дифференциация. 55 (1): 173–87. Дои:10.1111 / dgd.12020. PMID 23230908. S2CID 36706690.
- ^ Маунткасл V (1997). «Столбчатая организация неокортекса». Мозг. 120 (4): 701–722. Дои:10.1093 / мозг / 120.4.701. PMID 9153131.
- ^ Hubel DH, Wiesel TN (октябрь 1959 г.). «Рецептивные поля отдельных нейронов в полосатой коре головного мозга кошки». Журнал физиологии. 148 (3): 574–91. Дои:10.1113 / jphysiol.1959.sp006308. ЧВК 1363130. PMID 14403679.
- ^ С.М. Домбровски, К. Хильгетаг и Х. Барбас. Количественная архитектура отличает префронтальные корковые системы у макаки резус В архиве 2008-08-29 на Wayback Machine.Cereb. Кора 11: 975–988. «... они либо отсутствуют (агранулярный), либо имеют только рудиментарный зернистый слой IV (дисгранулярный)».
- ^ Сун В, Дэн И (2009). «Слоистые сетевые колебания и пространственно-временное рецептивное поле в зрительной коре». Proc Natl Acad Sci U S A. 106 (42): 17986–17991. Bibcode:2009PNAS..10617986S. Дои:10.1073 / pnas.0903962106. ЧВК 2764922. PMID 19805197.
- ^ Плетикос, Миховил; Соуза, Андре ММ; и другие. (22 января 2014 г.). "Временная спецификация и двусторонняя экспрессия топографических генов неокортекса человека". Нейрон. 81 (2): 321–332. Дои:10.1016 / j.neuron.2013.11.018. ЧВК 3931000. PMID 24373884.
- ^ Вольперт, Льюис (2015). Принципы развития (Пятое изд.). Великобритания: Издательство Оксфордского университета. п. 533. ISBN 9780199678143.
- ^ Уоррен Н., Карик Д., Пратт Т., Клаузен Дж. А., Асаваритикрай П., Мейсон Дж. О., Хилл Р. Э., Прайс-ди-джей (1999). «Фактор транскрипции Pax6 необходим для пролиферации и дифференцировки клеток в развивающейся коре головного мозга». Кора головного мозга. 9 (6): 627–35. Дои:10.1093 / cercor / 9.6.627. PMID 10498281.
- ^ Ларсен, У. Дж. Эмбриология человека, 3-е издание, 2001 г., стр. 421-422. ISBN 0-443-06583-7
- ^ Стивен К. Ноктор; Александр К. Флинт; Тамили А. Вайсман; Райан С. Даммерман и Арнольд Р. Кригштейн (2001). «Нейроны, происходящие из радиальных глиальных клеток, устанавливают радиальные единицы в неокортексе». Природа. 409 (6821): 714–720. Bibcode:2001Натура.409..714Н. Дои:10.1038/35055553. PMID 11217860. S2CID 3041502.
- ^ Сур, Мриганка; Лими, Кэтрин А. (2001). «Развитие и пластичность корковых областей и сетей». Обзоры природы Неврология. 2 (4): 251–262. Дои:10.1038/35067562. PMID 11283748. S2CID 893478.
- ^ а б c Sanes, Dan H .; Reh, Thomas A .; Харрис, Уильям А. (2012). Развитие нервной системы. Elsevier Inc. ISBN 978-0-12-374539-2.
- ^ Ракич, П. (октябрь 2009 г.). «Эволюция неокортекса: взгляд из биологии развития». Обзоры природы Неврология. 10 (10): 724–35. Дои:10.1038 / nrn2719. ЧВК 2913577. PMID 19763105.
- ^ Ракич, П. (ноябрь 1972 г.). «Внешние цитологические детерминанты дендритной структуры корзины и звездчатых клеток в молекулярном слое мозжечка». Журнал сравнительной неврологии. 146 (3): 335–54. Дои:10.1002 / cne.901460304. PMID 4628749. S2CID 31900267.
- ^ Гилберт, Скотт (2006). Биология развития (8-е изд.). Издательство Sinauer Associates. С. 394–395. ISBN 9780878932504.
- ^ Calegari, F; Haubensack W; Haffner C; Huttner WB (2005). «Избирательное удлинение клеточного цикла в нейрогенной субпопуляции нейральных клеток-предшественников во время развития мозга мышей». Журнал неврологии. 25 (28): 6533–8. Дои:10.1523 / jneurosci.0778-05.2005. ЧВК 6725437. PMID 16014714.
- ^ П. Ракич (1988). «Уточнение областей коры головного мозга». Наука. 241 (4862): 170–176. Bibcode:1988Научный ... 241..170R. Дои:10.1126 / science.3291116. PMID 3291116.
- ^ Hu, X.L .; Wang, Y .; Шен, К. (2012). «Эпигенетический контроль выбора клеточной судьбы в нервных стволовых клетках». Белки и клетки. 3 (4): 278–290. Дои:10.1007 / s13238-012-2916-6. ЧВК 4729703. PMID 22549586.
- ^ Костович, Ивица (1990). «История развития переходной зоны субпластинки в зрительной и соматосенсорной коре головного мозга макаки и человека». Журнал сравнительной неврологии. 297 (3): 441–470. Дои:10.1002 / cne.902970309. PMID 2398142. S2CID 21371568.
- ^ Ракич, П. (1 февраля 1974 г.). «Нейроны в зрительной коре головного мозга макаки-резуса: систематическая связь между временем происхождения и конечным расположением». Наука. 183 (4123): 425–7. Bibcode:1974Научный ... 183..425R. Дои:10.1126 / science.183.4123.425. PMID 4203022. S2CID 10881759.
- ^ Зечевич Н., Ракич П. (2001). «Развитие нейронов слоя I в коре головного мозга приматов». Журнал неврологии. 21 (15): 5607–19. Дои:10.1523 / JNEUROSCI.21-15-05607.2001. ЧВК 6762645. PMID 11466432.
- ^ Ракич, П. (8 июля 1988 г.). «Уточнение областей коры головного мозга». Наука. 241 (4862): 170–6. Bibcode:1988Научный ... 241..170R. Дои:10.1126 / science.3291116. PMID 3291116.
- ^ Фукути-Симогори, Т; Grove, EA (2 ноября 2001 г.). «Формирование паттерна неокортекса секретируемой сигнальной молекулой FGF8». Наука. 294 (5544): 1071–4. Bibcode:2001Научный ... 294.1071F. Дои:10.1126 / science.1064252. PMID 11567107. S2CID 14807054.
- ^ Гарель, С; Хаффман, KJ; Рубинштейн, JL (май 2003 г.). «Молекулярная регионализация неокортекса нарушена у гипоморфных мутантов Fgf8». Развитие. 130 (9): 1903–14. Дои:10.1242 / dev.00416. PMID 12642494.
- ^ Бишоп, км; Goudreau, G; О'Лири, Д. Д. (14 апреля 2000 г.). «Регулирование идентичности области в неокортексе млекопитающих с помощью Emx2 и Pax6». Наука. 288 (5464): 344–9. Bibcode:2000Sci ... 288..344B. Дои:10.1126 / science.288.5464.344. PMID 10764649.
- ^ Сыпь, БГ; Лим, HD; Breunig, JJ; Ваккарино, FM (26 октября 2011 г.). «Передача сигналов FGF увеличивает площадь поверхности коры эмбриона, регулируя Notch-зависимый нейрогенез». Журнал неврологии. 31 (43): 15604–17. Дои:10.1523 / jneurosci.4439-11.2011. ЧВК 3235689. PMID 22031906.
- ^ Раджагопалан, V; Скотт, Дж; Хабас, Пенсильвания; Ким, К; Corbett-Detig, J; Руссо, Ф; Баркович, AJ; Гленн, О.А.; Studholme, C (23 февраля 2011 г.). «Локальные модели роста тканей, лежащие в основе нормальной гирификации человеческого мозга плода, количественно определены в утробе матери». Журнал неврологии. 31 (8): 2878–87. Дои:10.1523 / jneurosci.5458-10.2011. ЧВК 3093305. PMID 21414909.
- ^ Луи, Ян Х .; Хансен, Дэвид V .; Кригштейн, Арнольд Р. (2011-07-08). «Развитие и эволюция неокортекса человека». Ячейка. 146 (1): 18–36. Дои:10.1016 / j.cell.2011.06.030. ISSN 1097-4172. ЧВК 3610574. PMID 21729779.
- ^ Шталь, Ронни; Уолчер, Тесса; Де Хуан Ромеро, Камино; Пильц, Грегор Александр; Каппелло, Сильвия; Ирмлер, Мартин; Санс-Аквела, Хосе Мигель; Бекерс, Йоханнес; Блюм, Роберт (25 апреля 2013 г.). «Trnp1 регулирует расширение и складывание коры головного мозга млекопитающих, контролируя судьбу радиальной глии». Ячейка. 153 (3): 535–549. Дои:10.1016 / j.cell.2013.03.027. ISSN 1097-4172. PMID 23622239.
- ^ Ван, Лэй; Хоу, Шируи; Хан, Ён Гу (2016-05-23). «Передача сигналов Hedgehog способствует расширению базальных предшественников, а также росту и сворачиванию неокортекса». Природа Неврология. 19 (7): 888–96. Дои:10.1038 / номер 4307. ISSN 1546-1726. ЧВК 4925239. PMID 27214567.
- ^ Сыпь, Брайан Дж .; Томази, Симона; Лим, Х. Дэвид; Suh, Carol Y .; Ваккарино, Флора М. (26.06.2013). «Гирификация коры, индуцированная фактором роста фибробластов 2 в мозге мышей». Журнал неврологии. 33 (26): 10802–10814. Дои:10.1523 / JNEUROSCI.3621-12.2013. ISSN 1529-2401. ЧВК 3693057. PMID 23804101.
- ^ Ракич, П. (8 июля 1988 г.). «Уточнение областей коры головного мозга». Наука. 241 (4862): 170–6. Bibcode:1988Научный ... 241..170R. Дои:10.1126 / science.3291116. PMID 3291116.
- ^ Фукути-Симогори, Т; Grove, EA (2 ноября 2001 г.). «Формирование паттерна неокортекса секретируемой сигнальной молекулой FGF8». Наука. 294 (5544): 1071–4. Bibcode:2001Научный ... 294.1071F. Дои:10.1126 / science.1064252. PMID 11567107. S2CID 14807054.
- ^ Бишоп, км; Goudreau, G; О'Лири, Д. Д. (14 апреля 2000 г.). «Регулирование идентичности области в неокортексе млекопитающих с помощью Emx2 и Pax6». Наука. 288 (5464): 344–9. Bibcode:2000Sci ... 288..344B. Дои:10.1126 / science.288.5464.344. PMID 10764649.
- ^ Grove, EA; Фукути-Симогори, Т. (2003). «Создание карты области коры головного мозга». Ежегодный обзор нейробиологии. 26: 355–80. Дои:10.1146 / annurev.neuro.26.041002.131137. PMID 14527269. S2CID 12282525.
- ^ Braitenberg, V и Schüz, A, 1998. "Cortex: Statistics and Geometry of Neuronal Connectivity. Второе тщательно переработанное издание" New York: Springer-Verlag
- ^ Саладин, Кеннет. Анатомия и физиология: единство формы и функции, 5-е изд. Нью-Йорк: McGraw-Hill Companies Inc., 2010. Печать.
- ^ Медицинский словарь Дорланда для потребителей здравоохранения, 2008 г.
- ^ Йео Б.Т., Криенен Ф.М., Сепулькр Дж., Сабунку М.Р., Лашкари Д., Холлинсхед М., Роффман Дж. Л., Смоллер Дж. В., Цёллей Л., Полимени Дж. Р., Фишл Б., Лю Х., Бакнер Р. Л. (2011). «Организация коры головного мозга человека оценивается по внутренним функциональным связям». Журнал нейрофизиологии. 106 (3): 1125–1165. Дои:10.1152 / ян.00338.2011. ЧВК 3174820. PMID 21653723.
- ^ Рупеш Кумар Шривастава; Юрген Шмидхубер (2014). «Понимание локально конкурентных сетей». arXiv:1410.1165 [cs.NE ].
- ^ Кэти Дж. Прайс (2000). «Анатомия языка: вклад функциональной нейровизуализации». Журнал анатомии. 197 (3): 335–359. Дои:10.1046 / j.1469-7580.2000.19730335.x. ЧВК 1468137. PMID 11117622.
- ^ Кентар, Модар; Манн, Мартина; Сам, Феликс; Оливарес-Ривера, Артуро; Санчес-Поррас, Ренан; Зереллес, Роланд; Саковиц, Оливер В .; Унтерберг, Андреас В .; Сантос, Эдгар (2020-01-15). «Обнаружение распространяющихся деполяризаций в модели окклюзии средней мозговой артерии у свиней». Acta Neurochirurgica. 162 (3): 581–592. Дои:10.1007 / s00701-019-04132-8. ISSN 0942-0940. PMID 31940093. S2CID 210196036.
- ^ Ortolano S, Vieitez I, et al. (2014). «Потеря корковых нейронов лежит в основе невропатологии болезни Лафора». Мол мозг. 7: 7. Дои:10.1186/1756-6606-7-7. ЧВК 3917365. PMID 24472629.
- ^ Mukherjee, Raja A.S .; Холлинз, Шейла (2006). "Расстройство алкогольного спектра плода: обзор". Журнал Королевского медицинского общества. 99 (6): 298–302. Дои:10.1258 / jrsm.99.6.298. ЧВК 1472723. PMID 16738372.
- ^ Тарапоре, ЧП; и другие. (Август 2012 г.). «Предоперационное мультимодальное моторное картирование: сравнение изображений магнитоэнцефалографии, управляемой транскраниальной магнитной стимуляции и прямой корковой стимуляции». Журнал нейрохирургии. 117 (2): 354–62. Дои:10.3171 / 2012.5.JNS112124. ЧВК 4060619. PMID 22702484.
- ^ а б c Уолш, Кристофер А .; Мочида, Ганешваран Х. (1 мая 2004 г.). «Генетические основы пороков развития коры головного мозга». Архив неврологии. 61 (5): 637–640. Дои:10.1001 / archneur.61.5.637. PMID 15148137.
- ^ "EMX2 пустые дыхальца гомеобокс 2 [Homo sapiens (человек)] - Ген - NCBI". www.ncbi.nlm.nih.gov.
- ^ Smigiel, R; Кабала, М; Якубяк, А; Кодера, H; Сасиадек, MJ; Мацумото, N; Сасиадек, ММ; Сайцу, Х (апрель 2016 г.). «Новая мутация COL4A1 у младенца с тяжелым дисморфическим синдромом с шизэнцефалией, перивентрикулярными кальцификациями и катарактой, напоминающей врожденную инфекцию». Исследование врожденных дефектов. Часть A, Клиническая и молекулярная тератология. 106 (4): 304–7. Дои:10.1002 / bdra.23488. PMID 26879631.
- ^ а б Принципы нейронауки. Кандел, Эрик Р. (5-е изд.). Нью-Йорк. 2013. С. 347–348. ISBN 9780071390118. OCLC 795553723.CS1 maint: другие (ссылка на сайт)
- ^ Томер, Р; Denes, AS; Тессмар-Райбл, К; Арендт, Д; Томер Р; Denes AS; Тессмар-Райбл К; Арендт Д. (2010). «Профилирование путем совмещения изображений показывает общее происхождение тел кольчатых грибов и паллиума позвоночных». Ячейка. 142 (5): 800–809. Дои:10.1016 / j.cell.2010.07.043. PMID 20813265. S2CID 917306.
внешние ссылки
- hier-20 в NeuroNames
- Изображения окрашенных срезов головного мозга, которые включают «кору головного мозга» на Проект BrainMaps
- «Первичная зрительная кора», Webvision: Подробная статья о структуре и функциях первичной зрительной коры.
- «Основные типы клеток», Webvision: изображение основных типов клеток коры головного мозга обезьян.
- Кора головного мозга - база данных, ориентированная на клетки
{kind=link}