Коинофилия - Koinophilia


Коинофилия является эволюционный гипотеза, предполагающая, что во время половой отбор, животные преимущественно ищут товарищи с минимумом необычных или мутант особенности, включая функциональность, внешний вид и поведение.[1][2][3][4][5][6] Койнофилия пытается объяснить кластеризацию половых организмов в разновидность и другие проблемы, описанные Дилемма Дарвина.[3][4][5] Термин происходит от греческого: коино, «общий», «общий» и филия, "нежность".
Естественный отбор причины выгодный унаследованные функции становятся более распространенными за счет их невыгодных аналогов. Гипотеза койнофилии предполагает, что от репродуктивного животного, следовательно, можно ожидать, что он будет избегать особей с редкими или необычными особенностями и предпочитать спариваться с особями, демонстрирующими преобладание общих или средних признаков.[2][3] Мутанты со странными, странными или необычными характеристиками можно было бы избежать, потому что большинство мутаций, которые проявляются в изменении внешнего вида, функциональности или поведения, являются невыгодными.[7] Поскольку невозможно судить о том, является ли новая мутация полезной (или может быть выгодна в непредвиденном будущем) или нет, койнофильные животные избегают их всех за счет избежания очень случайных потенциально полезных мутаций.[8] Таким образом, койнофилия, хотя и не безупречная в своей способности различать поместиться от неподходящих товарищей, хорошая стратегия, когда выбор супруга. Койнофильный выбор гарантирует, что потомство, вероятно, унаследует набор функций и атрибутов, которые хорошо служили всем членам вида в прошлом.[3]
Койнофилия отличается от модели спаривания типа «подобное предпочитает подобное». ассортативная вязка.[9][10] Если нравится, нравится, лейкистический животные (например, белые павлины) будут испытывать сексуальное влечение друг к другу, и возникнет лейкистический подвид. Койнофилия предсказывает, что это маловероятно, потому что лейцистические животные привлекаются к среднему так же, как и все другие представители этого вида. Поскольку нелейцистические животные не привлекаются лейцизмом, немногие лейцистические особи находят себе пару, и лейцистические линии образуются редко.
Койнофилия дает простые объяснения почти универсальным канализация сексуальных существ на виды,[3][4][5] редкость переходных форм между видами (как между современными, так и ископаемыми видами),[3][4] эволюционный застой, прерывистое равновесие,[3][4][5] и эволюция сотрудничество.[11][12] Койнофилия также может способствовать поддержание из половое размножение, предотвращая его возврат к более простому бесполая форма размножения.[13][14]
Гипотеза койнофилии подтверждается открытиями Джудит Ланглуа и ее коллег.[2][15][16][17] Они обнаружили, что среднее из двух человеческих лиц было более привлекательным, чем любое из лиц, из которых это среднее было получено.[18] Чем больше лиц (одного пола и возраста) использовалось в процесс усреднения тем более привлекательным и привлекательным становилось среднее лицо.[19] Эта работа в усредненность[2][15][16][20] поддерживает койнофилию как объяснение того, что составляет красивая лицо.[17][21][22]
Видообразование и прерывистые равновесия
Биологи от Дарвина и далее [23] ломали голову над тем, как эволюция производит виды чьи взрослые особи необычайно похожи и сильно отличаются от представителей других видов. Львы и леопарды являются, например, большими плотоядными животными, обитающими в одной и той же окружающей среде, и охотятся на одну и ту же добычу, но выглядят совершенно по-разному. Вопрос в том, почему не существует промежуточных звеньев.[7][24]

Это «горизонтальное» измерение двумерной задачи,[28][29] имеется в виду почти полное отсутствие переходных или промежуточных форм между современными видами (например, между львами, леопардами и гепардами).[24][30][31]
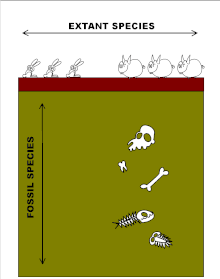
«Вертикальное» измерение касается летописи окаменелостей. Ископаемые виды часто удивительно стабильны в течение чрезвычайно длительных геологических периодов, несмотря на дрейф континентов, серьезные изменения климата и массовые вымирания.[32][33][34] Когда происходит изменение формы, оно имеет тенденцию быть резким в геологическом плане, снова вызывая фенотипические разрывы (то есть отсутствие промежуточных форм), но теперь между последовательными видами, которые затем часто сосуществуют в течение длительных периодов времени. Таким образом, летопись окаменелостей предполагает, что эволюция происходит всплесками, перемежающимися длительными периодами эволюционной стагнации в так называемых прерывистое равновесие.[32] Почему это так, с тех пор остается загадкой эволюции. Дарвин первым осознал проблему.[23][34][35]
Койнофилия может объяснить как горизонтальные, так и вертикальные проявления видообразование и почему он, как правило, затрагивает весь внешний вид соответствующих животных.[3][4][5] Поскольку койнофилия влияет на весь По внешнему виду члены скрещивающейся группы стремятся выглядеть одинаково во всех деталях.[25][36] Каждая межпородная группа быстро разовьет свой характерный внешний вид.[5] Следовательно, особь из одной группы, которая переходит в другую, будет признана другой и будет подвергаться дискриминации в течение брачного сезона. Репродуктивная изоляция, вызванная койнофилией, может, таким образом, стать первым решающим шагом в развитии, в конечном счете, физиологических, анатомических и поведенческих барьеров для гибридизации и, таким образом, в конечном итоге, полного видового развития. Впоследствии койнофилия будет защищать внешний вид и поведение этого вида от вторжения необычных или незнакомых форм (которые могут возникнуть в результате иммиграции или мутации) и, таким образом, станет парадигмой прерывистое равновесие (или «вертикальный» аспект проблемы видообразования).[3][4]
Эволюция в койнофильных условиях








Фон
Эволюция может быть чрезвычайно быстрой, о чем свидетельствует создание одомашненных животных и растений за очень короткий геологический период, охватывающий всего несколько десятков тысяч лет, людьми, почти или совсем не знающими генетику. Кукуруза, Zea Mays, например, был созданный в Мексике всего за несколько тысяч лет, примерно от 7 000 до 12 000 лет назад.[37] Это поднимает вопрос, почему в долгосрочной перспективе скорость эволюции намного медленнее, чем теоретически возможно.[7][32][34][38]
Эволюция навязывается видам или группам. В некоторых странах это не планируется и не предпринимается. Ламаркист путь.[39][40] Мутации, от которых зависит процесс, являются случайными событиями, и, за исключением "тихие мутации "которые не влияют на функциональность или внешний вид носителя, поэтому обычно являются невыгодными, и их шансы оказаться полезными в будущем исчезающе малы. Следовательно, хотя вид или группа могут выиграть, имея возможность адаптироваться к новому окружающей среде за счет накопления широкого диапазона генетических вариаций, это в ущерб отдельные лица которые должны нести эти мутации до тех пор, пока небольшое, непредсказуемое меньшинство из них в конечном итоге не внесет вклад в такую адаптацию. Таким образом способность развиваться - это групповая адаптация, который был дискредитирован, среди прочего, Джордж К. Уильямс,[41] Джон Мейнард Смит[42] и Ричард Докинз.[43][44][45][46] потому что это не на пользу человеку.
Следовательно, сексуальная отдельные лица можно ожидать, что они будут избегать передачи мутаций своему потомству, избегая самок со странными или необычными характеристиками.[1][2][3][5] Мутации, которые поэтому влияют на внешний вид и привычки их носителей, редко передаются следующему и последующим поколениям. Поэтому они редко будут проверяться естественным отбором. Таким образом, эволюционные изменения в большой популяции с широким выбором партнеров практически остановятся. Единственные мутации, которые могут накапливаться в популяции, - это мутации, не оказывающие заметного влияния на внешний вид и функциональность их носителей (их, таким образом, называют "тихий " или же "нейтральные мутации ").
Эволюционный процесс
Сдержанность койнофилии оказывает на фенотипическое изменение предполагает, что эволюция может происходить только в том случае, если мутантных партнеров нельзя избежать в результате острой нехватки потенциальных партнеров. Скорее всего, это произойдет в небольшие ограниченные сообщества, например, на небольших островах, в отдаленных долинах, озерах, речных системах, пещерах,[9] или же в периоды оледенения,[47] или после массовые вымирания, когда можно ожидать внезапных всплесков эволюции.[48] В этих условиях не только сильно ограничивается выбор партнеров, но и узкие места населения, эффекты основателя, генетический дрейф и инбридинг вызывают быстрые, случайные изменения в генетическом составе изолированной популяции.[9] Более того, гибридизация с родственными видами заключенные в тот же изолят, могут привести к дополнительным генетическим изменениям.[49][50] Если изолированное население, такое как эта переживает свои генетические потрясения, и впоследствии расширится в незанятую нишу или в нишу, в которой он имеет преимущество перед своими конкурентами, возникнет новый вид или подвид. С геологической точки зрения это будет резкое событие. После этого возобновление избегания мутантных партнеров снова приведет к эволюционной стагнации.
Таким образом Окаменелости эволюционный прогресс обычно состоит из видов, которые внезапно появляются и в конечном итоге исчезают через сотни тысяч или миллионы лет без каких-либо изменений внешнего вида.[33][35][48][51] Графически эти ископаемые виды представлены горизонтальными линиями, длина которых показывает, как долго каждый из них существовал. Горизонтальность линий показывает неизменный внешний вид каждого ископаемого вида, изображенного на графике. Во время существования каждого вида новые виды появляются через случайные промежутки времени, каждый также существует много сотен тысяч лет, прежде чем исчезнуть без изменения внешнего вида. Степень родства и линии происхождения этих параллельных видов, как правило, невозможно определить. Это показано на следующей схеме, изображающей эволюция современного человека с тех пор, как гоминины отделенные от линии, которая привела к эволюции наших ближайших ныне живущих родственников приматов, шимпанзе.[51]

Фенотипические последствия
Это предложение, что узкие места населения возможно, являются основными генераторами вариаций, которые подпитывают эволюцию, предсказывает, что эволюция обычно будет происходить в прерывистом, относительно большом масштабе. морфологический шаги, перемежающиеся длительными периодами эволюционного застоя,[52] вместо непрерывной серии мелко дифференцированных изменений.[53] Однако он делает дальнейший прогноз.[4] Дарвин подчеркнул, что общие биологически бесполезные странности и несоответствия, которые характеризуют вид, являются признаками эволюционной истории - чего нельзя было бы ожидать, если бы, например, крыло птицы было спроектировано de novo, как утверждали его недоброжелатели.[54] Настоящая модель предсказывает, что, помимо пережитков, отражающих эволюционное наследие организма, все представители данного вида также будут нести печать своего изоляционного прошлого - произвольные, случайные особенности, накопленные в результате эффекты основателя, генетический дрейф и другие генетические последствия полового размножения у небольшие изолированные сообщества.[4][55] Таким образом, все львы, африканские и азиатские, имеют очень характерный черный пучок меха на конце хвоста, который трудно объяснить с точки зрения адаптации или как следы раннего кошачий, или более древний предок. Уникальное, часто богатое цветами и узорами оперение каждого из самых разнообразных современных видов птиц представляет собой аналогичную загадку эволюции. Этот разнообразный набор фенотипы легче объяснить как продукты изолятов, впоследствии защищаемых койнофилией, чем как совокупность очень разнообразных эволюционных реликвий или как совокупность уникально эволюционирующих приспособлений.

Черный пучок меха в конце этого Азиатский лев хвост трудно объяснить как эволюционный пережиток или адаптацию. Тем не менее, это происходит у всех львов.

«Очки» очковый буревестник, Procellaria conspicillata, представляют собой необъяснимую особенность этого вида.

Уникальное оперение обыкновенный зяблик, Fringilla coelebs, безошибочно идентифицирует вид, но его адаптивная роль загадочна.



Эволюция сотрудничества
Сотрудничество - это любое групповое поведение, которое приносит больше пользы людям, чем если бы они действовали как независимые агенты.

Однако эгоистичные люди могут использовать сотрудничество других, не принимая участия в групповой деятельности, но все же пользуясь ее преимуществами. Например, эгоистичный человек, который не присоединяется к охотничьей стае и не разделяет ее риски, но, тем не менее, разделяет добычу, имеет преимущество в пригодности перед другими членами стаи. Таким образом, хотя группа кооперативных индивидуумов более приспособлена, чем эквивалентная группа эгоистичных индивидуумов, эгоистичные индивидуумы, вкрапленные в сообщество кооператоров, всегда более приспособлены, чем их хозяева. В среднем они производят больше потомства, чем их хозяева, и в конечном итоге заменят их.[43][44][45][46]
Если, однако, эгоистичные люди подвергаются остракизму и отвергаются как партнеры из-за их девиантного и необычного поведения, то их эволюционное преимущество становится эволюционной помехой.[3] Тогда сотрудничество становится эволюционно стабильный.[11][12]
Влияние диет и условий окружающей среды

Наиболее документированные создания новых видов в лаборатории были выполнены в конце 1980-х годов. Уильям Райс и Г. Соленые плодовые мухи, Drosophila melanogaster, используя лабиринт с тремя различными вариантами среды обитания, такими как светлый / темный и влажный / сухой. Каждое поколение было помещено в лабиринт, и группы мух, вышедших из двух из восьми выходов, были разделены для размножения друг с другом в своих группах. После тридцати пяти поколений две группы и их потомство были репродуктивно изолированы из-за их сильных предпочтений в среде обитания: они спаривались только в тех областях, которые они предпочитали, и поэтому не спаривались с мухами, которые предпочитали другие области.[56] История таких попыток описана в Rice and Hostert (1993).[57][58]
Дайан Додд использовала лабораторный эксперимент, чтобы показать, как репродуктивная изоляция может развиваться в Drosophila pseudoobscura плодовых мушек через несколько поколений, поместив их в различные среды, на основе крахмала или мальтозы.[59]

Эксперимент Додда легко повторить и многим другим, в том числе с другими видами плодовых мушек и другими продуктами питания.[60]
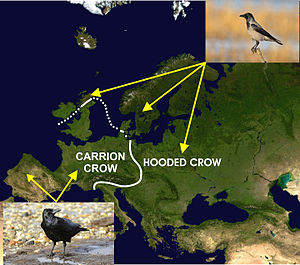
В падальщик (Corvus corone) и ворон (Corvus Cornix) - два близкородственных вида, географическое распространение которых в Европе показано на прилагаемой диаграмме. Считается, что это распространение могло быть результатом циклов оледенения во время Плейстоцен, что привело к разделению родительской популяции на изоляты, которые впоследствии снова расширили свои ареалы при потеплении климата, что привело к вторичному контакту.[47][61] Джелмер В. Поэльстра и его коллеги секвенировали почти все геномы обоих видов в популяциях на разном расстоянии от зоны контакта, чтобы обнаружить, что эти два вида генетически идентичны как по своей ДНК, так и по экспрессии (в форме РНК), за исключением для отсутствие выражения небольшой части (<0,28%) генома (расположенной на птичьей хромосоме 18) ворон с капюшоном, что придает более светлую окраску оперения ее туловищу.[47] Таким образом, эти два вида могут жизнеспособно гибридизироваться, и иногда это происходит в зоне контакта, но полностью черные вороны-падальщики на одной стороне зоны контакта спариваются почти исключительно с другими полностью черными воронами-падальщиками, в то время как то же самое происходит среди черных воронов. вороны по ту сторону зоны контакта. Поэтому ясно, что только внешний вид двух видов препятствует гибридизации.[47][61] Авторы связывают это с ассортативная вязка, преимущество которого неясно, и это привело бы к быстрому появлению потоков новых линий и, возможно, даже видов из-за взаимного притяжения между мутантами. Унникришнан и Ахила[62] предложить вместо этого, что койнофилия - это более точный объяснение устойчивости к гибридизации в зоне контакта, несмотря на отсутствие физиологических, анатомических или генетических барьеров для такой гибридизации.
Прием
Уильям Б. Миллер,[5] в недавнем обширном обзоре теории коинофилии (2013 г.) отмечает, что точный Объяснение группировки половых животных по видам, их неизменной устойчивости в летописи окаменелостей в течение длительного периода времени и фенотипических разрывов между видами, как ископаемыми, так и дошедшими до нас, представляет собой серьезный отход от широко принятой точки зрения полезные мутации распространяться, в конечном счете, на всю или некоторую часть популяции (заставляя ее эволюционировать ген за геном).[63][64] Дарвин признал, что этот процесс не имел врожденной или неизбежной склонности к воспроизводству видов.[24][23] Вместо этого население будет в вечное состояние перехода как во времени, так и в пространстве.[24][23] В любой данный момент они будут состоять из людей с различным числом полезных характеристик, которые могли или не могли достичь их из различных точек происхождения в популяции, а нейтральные черты будут иметь разброс, определяемый случайными механизмами, такими как генетический дрейф.[65][66][67]
Он также отмечает, что койнофилия не объясняет, как возникают физиологические, анатомические и генетические причины репродуктивной изоляции. Это только поведенческий репродуктивная изоляция к которому обращается койнофилия. Более того, трудно понять, как койнофилия может применяться к растениям и определенным морским существам, которые выбрасывают свои гаметы в окружающую среду для встречи и слияния, по-видимому, совершенно случайным образом (в пределах определенного вида). Однако, когда для опыления стигмат используется пыльца от нескольких совместимых доноров, доноры обычно не дают равное количество семян.[68] Маршалл и Диггл заявляют, что существование некоторого неслучайного отцовства семян у цветковых растений на самом деле не обсуждается. Как это происходит, остается неизвестным. Выбор пыльцы - одна из возможностей,[68] с учетом того, что 50% гаплоидного генома пыльцевого зерна экспрессируется при росте его трубки к семяпочке.[69]
Явное предпочтение самок некоторых, особенно птиц, видов преувеличенные мужские украшения, например, хвост павлина,[7][70][71] нелегко примирить с концепцией койнофилии.
Рекомендации
- ^ а б Саймонс, Дональд (1979). Эволюция человеческой сексуальности. Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-502535-4. LCCN 78023361. OCLC 4494283.
- ^ а б c d е Langlois, Judith H .; Роггман, Лори А. (1990). «Привлекательные лица только средние». Психологическая наука. 1 (2): 115–121. Дои:10.1111 / j.1467-9280.1990.tb00079.x. ISSN 0956-7976. S2CID 18557871.
- ^ а б c d е ж грамм час я j k Коэслаг, Йохан Х. (1990). «Койнофилия группирует сексуальных существ по видам, способствует застою и стабилизирует социальное поведение». Журнал теоретической биологии. 144 (1): 15–35. Дои:10.1016 / s0022-5193 (05) 80297-8. ISSN 0022-5193. PMID 2200930.
- ^ а б c d е ж грамм час я Коэслаг, Йохан Х. (1995). «О двигателе видообразования». Журнал теоретической биологии. 177 (4): 401–409. Дои:10.1006 / jtbi.1995.0256. ISSN 0022-5193.
- ^ а б c d е ж грамм час Миллер, Уильям Б. (2013). «Что такого особенного в эволюционных пробелах?». В: Микрокосм внутри: Эволюция и исчезновение в гологеноме. Бока-Ратон, Флорида: Universal Publishers. С. 177, 395–396. ISBN 978-1-61233-2772.
- ^ Унникришнан, М. (2012). «Койнофилия снова: эволюционная связь между выбором партнера и распознаванием лиц». Текущая наука. 102 (4): 563–570.
- ^ а б c d е Мэйнард Смит, Джон (1989). Эволюционная генетика. Оксфорд: Издательство Оксфордского университета. С. 55, 267, 273, 281. ISBN 978-0-19-854215-5.
- ^ Уильямс, Джордж К. (1992). «Оптимизация и связанные с ней концепции». В: Естественный отбор: области, уровни и проблемы. Нью-Йорк: Издательство Оксфордского университета. С. 60–61. ISBN 978-0-19-506933-4.
- ^ а б c Айяла, Франсиско Дж. (1982). Популяционная и эволюционная генетика. Менло-Парк, Калифорния: Бенджамин / Каммингс. стр.60, 73–83, 182–190, 198–215. ISBN 978-0-8053-0315-5. LCCN 81021623. OCLC 8034790.
- ^ Бусс, Д. (1985). «Выбор спутника жизни». Американский ученый. 37 (1): 47–51. Bibcode:1985AmSci..73 ... 47B.
- ^ а б Koeslag, J.H. (1997). «Секс, дилемма заключенного и эволюционная неизбежность сотрудничества». Журнал теоретической биологии. 189 (1): 53–61. Дои:10.1006 / jtbi.1997.0496. PMID 9398503.
- ^ а б Koeslag, J.H. (2003). «Эволюция сотрудничества: сотрудничество побеждает дезертирство в модели кукурузного поля». Журнал теоретической биологии. 224 (3): 399–410. Дои:10.1016 / s0022-5193 (03) 00188-7. PMID 12941597.
- ^ Koeslag, J.H .; Коеслаг, П. (1993). «Эволюционно устойчивый мейотический пол». Журнал наследственности. 84 (5): 396–399. Дои:10.1093 / oxfordjournals.jhered.a111360.
- ^ Koeslag, P.D .; Koeslag, J.H. (1994). «Койнофилия стабилизирует двухполое половое размножение против бесполого в неизменной среде». Журнал теоретической биологии. 166 (3): 251–260. Дои:10.1006 / jtbi.1994.1023. PMID 8159013.
- ^ а б Langlois, J.H .; Roggman, L.A .; Musselman, L .; Актон, С. (1991). "Картинка лучше тысячи слов: ответ на вопрос 'О сложности усреднения лиц'". Психологическая наука. 2 (5): 354–357. Дои:10.1111 / j.1467-9280.1991.tb00165.x. S2CID 143993972.
- ^ а б Langlois, J.H .; Массельман, Л. (1995). «Мифы и загадки красоты». В Калхуне, Д. Р. (ред.). Ежегодник науки и будущего 1996 г.. Чикаго: Британская энциклопедия, Inc., стр. 40–61.
- ^ а б Rubenstein, A.J .; Langlois, J.H .; Roggman, J.H. (2002). «Что делает лицо привлекательным и почему: роль среднего в определении красоты лица». В Родосе, G .; Зебровиц, Л.А. (ред.). Внешняя привлекательность: эволюционная, когнитивная и социальная точки зрения. Вестпорт, Коннектикут: Ablex. С. 1–33.
- ^ Мартин Грюндль. "среднее двух человеческих лиц". Uni-regensburg.de. Получено 2018-06-04.
- ^ «процесс усреднения тем более привлекательным и привлекательным становилось среднее лицо». Homepage.psy.utexas.edu. Архивировано из оригинал на 2015-02-04. Получено 2018-06-04.
- ^ Langlois, J.H .; Roggman, L.A .; Массельман, Л. (1994). «Что является средним, а что нет в привлекательных лицах?». Психологическая наука. 5 (4): 214–220. Дои:10.1111 / j.1467-9280.1994.tb00503.x. S2CID 145147905.
- ^ Калик, S.M .; Zebrowitz, L.A .; Langlois, J.H .; Джонсон, Р. (1998). «Действительно ли привлекательность человеческого лица рекламирует здоровье? Продольные данные по эволюционному вопросу». Психологическая наука. 9: 8–13. Дои:10.1111/1467-9280.00002. S2CID 9134500.
- ^ Hoss, R.A .; Ланглуа, Дж. (2003). «Младенцы предпочитают мордашки». В Pascalis, O .; Слейтер, А. (ред.). Развитие обработки лица в младенчестве и раннем детстве: современные перспективы. Нью-Йорк: Nova Science Publishers. С. 27–38.
- ^ а б c d Дарвин, Чарльз (1859 г.). «Глава 6,« Трудности теории »в». О происхождении видов путем естественного отбора или о сохранении избранных рас в борьбе за жизнь (1-е изд.). Лондон: Джон Мюррей. LCCN 06017473. OCLC 741260650.
- ^ а б c d е Гулд, Стивен Джей (1980). «Куахог - это куахог». В: Большой палец панды: больше размышлений в естественной истории. Нью-Йорк: W.W. Нортон и компания. стр.204–213. ISBN 978-0-393-30023-9.
- ^ а б Хоккей, P.A.R .; Dean, W.R.J .; Райан, П.Г., ред. (2003). Робертс - Птицы Южной Африки, VII изд.. Кейптаун: Птичий фонд Джона Фолькера. С. 14, 60–1138. ISBN 978-0620340519.
- ^ White, G.C .; Бернхэм, К. (1999). «Программа МАРК. Оценка выживаемости помеченных животных». Исследование птиц. 46: S120 – S139. Дои:10.1080/00063659909477239.
- ^ Рейнольдс, Ричард Т .; Джой, Сюзанна М .; Лесли, Дуглас Г. (1994).«Гнездо продуктивности, верности и расстояния между ястребами-тетеревятниками в Аризоне». Исследования по птичьей биологии. 16: 106–113.
- ^ Бернштейн, Харрис; Байерли, Генри К.; Hopf, Frederic A .; и другие. (1985). «Секс и появление видов». Журнал теоретической биологии. 117 (4): 665–690. Дои:10.1016 / S0022-5193 (85) 80246-0. ISSN 0022-5193. PMID 4094459.
- ^ Хопф, Фредерик А. (1990). «Дилемма Дарвина переходных форм: сравнение модели с данными». В Maynard Smith, J .; Вида, Г. (ред.). Организационные ограничения динамики эволюции. Манчестер: Издательство Манчестерского университета. С. 357–372.
- ^ Гулд, Стивен Джей (1985). Улыбка фламинго: размышления в естествознании. Лондон: Penguin Books. С. 160, 169. ISBN 978-0-14-013554-1.
- ^ Мейнард Смит, Джон (1983). «Генетика застоя и пунктуации». Ежегодный обзор генетики. 17: 11–25. Дои:10.1146 / annurev.ge.17.120183.000303. ISSN 0066-4197. PMID 6364957.
- ^ а б c Элдридж, Найлз; Гулд, Стивен Джей (1972). «Прерывистое равновесие: альтернатива филетической постепенности». В Schopf, T.J.M. (ред.). Модели в палеобиологии. Сан-Франциско: Фриман Купер. С. 82–115.
- ^ а б Гулд, Стивен Джей; Элдридж, Найлз (1977). «Прерывистые равновесия: пересмотр темпов и способов эволюции». Палеобиология. 3 (2): 115–151. Дои:10.1017 / S0094837300005224.
- ^ а б c Уильямс, Джордж К. (1992). «Стазис». В: Естественный отбор: области, уровни и проблемы. Нью-Йорк: Издательство Оксфордского университета. С. 127–142. ISBN 978-0-19-506933-4.
- ^ а б Гулд, Стивен Джей (1980). «Эпизодичность эволюционных изменений». В: Большой палец панды: больше размышлений в естественной истории. Нью-Йорк: W.W. Нортон и компания. стр.179–185. ISBN 978-0-393-30023-9.
- ^ Стюарт, Крис; Стюарт, Тильда (2007). Полевой справочник по млекопитающим юга Африки (Четвертое изд.). Кейптаун: Струик. С. 30–285. ISBN 978-1-77007-404-0.
- ^ Законы, Билл (2010). Пятьдесят растений, изменивших ход истории. Буффало, Нью-Йорк: Книги Светлячка. С. 210–215. ISBN 978-1-55407-798-4. LCCN 2011414731. OCLC 711609823.
- ^ Майр, Эрнст (1954). «Изменение генетической среды и эволюция». В Хаксли, Джулиан; Харди, Алистер С.; Форд, Эдмунд Б. (ред.). Эволюция как процесс. Лондон: Аллен и Анвин. LCCN 54001781. OCLC 974739.
- ^ Гулд, Стивен Джей (1980). «Возвращение обнадеживающего монстра». В: Большой палец панды: больше размышлений в естественной истории. Нью-Йорк: W.W. Нортон и компания. стр.186–193. ISBN 978-0-393-30023-9.
- ^ Докинз, Ричард (1988). Слепой часовщик. Лондон: Penguin Books. С. 5, 21. ISBN 978-0-14-014481-9.
- ^ Уильямс, Джордж К. (1972). Адаптация и естественный отбор: критика некоторых современных эволюционных идей. Принстон: Издательство Принстонского университета. ISBN 978-0-691-02357-1.
- ^ Мейнард Смит, Джон (1964). «Групповой отбор и родовой отбор». Природа. 201 (4924): 1145–1147. Bibcode:1964Натура.201.1145S. Дои:10.1038 / 2011145a0. S2CID 4177102.
- ^ а б Докинз, Ричард (1995). Река вне Эдема: дарвиновский взгляд на жизнь. Серия «Мастера науки». Нью-Йорк: Базовые книги. ISBN 978-0-465-01606-8. LCCN 94037146. OCLC 31376584.
- ^ а б Докинз, Р. (1994). "Похоронение автомобиля Комментарий к Wilson & Sober: Выбор группы". Поведенческие науки и науки о мозге. 17 (4): 616–617. Дои:10.1017 / S0140525X00036207. Архивировано из оригинал 15 сентября 2006 г.
- ^ а б Деннет, Дэниел С. (1994). "E Pluribus Unum? Комментарий к Wilson & Sober: Group Selection". Поведенческие науки и науки о мозге. 17 (4): 617–618. Дои:10.1017 / с0140525x00036219. Архивировано из оригинал 27 декабря 2007 г.
- ^ а б Пинкер, С. (2012). Ложное очарование группового отбора. Edge, 19 июня 2012 г. http://edge.org/conversation/the-false-allure-of-group-selection
- ^ а б c d Poelstra, Jelmer W .; Виджай, Нагарджун; Bossu, Christen M .; и другие. (2014). «Геномный ландшафт, лежащий в основе фенотипической целостности перед лицом потока генов у ворон». Наука. 344 (6190): 1410–1414. Bibcode:2014Научный ... 344.1410P. Дои:10.1126 / science.1253226. ISSN 0036-8075. PMID 24948738. S2CID 14431499.
- ^ а б Кэмпбелл, Нил А. (1990). Биология (2-е изд.). Редвуд-Сити, Калифорния: Паб Бенджамин / Каммингс. Co., стр. 450–451, 487–490, 499–501. ISBN 978-0-8053-1800-5. LCCN 89017952. OCLC 20352649.
- ^ Арнольд, М. (1996). Естественная гибридизация и эволюция. Нью-Йорк: Издательство Оксфордского университета. п. 232. ISBN 978-0-19-509975-1.
- ^ Wendel, J.F .; Дойл, Дж. Дж. (1998). «Секвенирование ДНК». In Soltis, D.E .; Солтис, П.С.; Дойл, Дж. Дж. (ред.). Молекулярная систематика растений II. Бостон: Клувер. С. 265–296.
- ^ а б Маккарти, Теренс; Рубидж, Брюс (2005). История Земли и Жизни: взгляд на юг Африки на 4,6 миллиарда лет путешествия. Кейптаун, Южная Африка: Struik Publishers. ISBN 978-1-77007-148-3. LCCN 2006376206. OCLC 62098231.
- ^ Гулд, Стивен Джей (1980). «Эпизодичность эволюционных изменений». В: Большой палец панды: больше размышлений в естественной истории. Нью-Йорк: W.W. Нортон и компания. стр.179–185. ISBN 978-0-393-30023-9.
- ^ Докинз, Ричард (1988). «Прокладывая следы в пространстве животных» В: Слепой часовщик. Лондон: Penguin Books. С. 77–109. ISBN 978-0-14-014481-9.
- ^ Гулд, Стивен Джей (1980). «Бессмысленные знаки истории». В: Большой палец панды: больше размышлений в естественной истории. Нью-Йорк: W.W. Нортон и компания. стр.27–34. ISBN 978-0-393-30023-9.
- ^ Циммер, Карл (2001). Эволюция: торжество идеи. Введение Стивен Джей Гулд; предисловие Ричарда Хаттона (1-е изд.). Нью-Йорк: ХарперКоллинз. ISBN 978-0-06-019906-7. LCCN 2001024077. OCLC 46359440.
- ^ Райс, Уильям Р .; Соль, Джордж У. (1988). «Видообразование посредством разрушающего отбора по предпочтениям среды обитания: экспериментальные данные». Американский натуралист. 131 (6): 911–917. Дои:10.1086/284831. ISSN 0003-0147.
- ^ Райс, Уильям Р .; Хостерт, Эллен Э. (1993). «Лабораторные эксперименты по видообразованию: что мы узнали за 40 лет?». Эволюция. 47 (6): 1637–1653. Дои:10.2307/2410209. ISSN 0014-3820. JSTOR 2410209. PMID 28568007.
- ^ Гаврилец, Сергей (2003). «Перспектива: модели видообразования: что мы узнали за 40 лет?». Эволюция. 57 (10): 2197–2215. Дои:10.1554/02-727. ISSN 0014-3820. PMID 14628909. S2CID 198158082.
- ^ Додд, Дайан М. Б. (1989). "Репродуктивная изоляция как следствие адаптивной дивергенции в Drosophila pseudoobscura". Эволюция. 43 (6): 1308–1311. Дои:10.2307/2409365. ISSN 0014-3820. JSTOR 2409365. PMID 28564510.
- ^ Киркпатрик, Марк; Равинье, Вирджиния (2002). «Видообразование путем естественного и полового отбора: модели и эксперименты». Американский натуралист. 159 (S3): S22 – S35. Дои:10.1086/338370. ISSN 0003-0147. PMID 18707367.
- ^ а б де Книжф, Питер (2014). «Как падаль и вороны побеждают проклятие Линнея». Наука. 344 (6190): 1345–1346. Bibcode:2014Научный ... 344.1345D. Дои:10.1126 / science.1255744. ISSN 0036-8075. PMID 24948724. S2CID 207790306.
- ^ Poelstra, Jelmer W .; Виджай, Нагарджун; Bossu, Christen M .; и другие. (2014). «Геномный ландшафт, лежащий в основе фенотипической целостности перед лицом потока генов у ворон» (PDF). Наука. 344 (6190): 1410–1414. Bibcode:2014Научный ... 344.1410P. Дои:10.1126 / science.1253226. ISSN 0036-8075. PMID 24948738. S2CID 14431499.
Фенотипические различия между падалью и серой вороной в зоне гибридизации в Европе вряд ли связаны с ассортативным скрещиванием.
- Комментарий Мажуванчерри К. Унникришнана и Х. С. Ахилы. - ^ Докинз, Ричард (1988). «Накопление мелочи». В: Слепой часовщик. Лондон: Penguin Books. С. 43–74. ISBN 978-0-14-014481-9.
- ^ Докинз, Ричард (1996). Восхождение на маунт Невероятно. Лондон: Penguin Books. С. 97–179. ISBN 978-0-670-85018-1.
- ^ Gould, S.J .; Левонтин, Р. (Сентябрь 1979 г.). «Spandrels Сан-Марко и парадигмы Panglossian: критика адаптационистской программы». Труды Лондонского королевского общества B (опубликовано в 1979 г.). 205 (1161): 581–598. Bibcode:1979RSPSB.205..581G. Дои:10.1098 / rspb.1979.0086. PMID 42062. S2CID 2129408.
- ^ Гулд, Стивен Джей (1983). «Мифы и реалии гиены». В: Куриные зубы и конские пальцы. Дальнейшие размышления в естественной истории. Хармондсворт, Мидлсекс, Англия: Penguin Books. стр.147–157. ISBN 978-0-14-022533-4.
- ^ Гулд, Стивен Джей (1980). Большой палец панды. Больше размышлений в естественной истории. Нью-Йорк: W.W. Нортон и компания. стр.129–130. ISBN 978-0-393-30023-9.
- ^ а б Маршалл, Дайан Л .; Диггл, Памела К. (2001). «Механизмы дифференциальной донорской активности пыльцы дикого редиса. Raphanus sativus (Brassicaceae) ". Американский журнал ботаники. 88 (2): 242–257. Дои:10.2307/2657015. ISSN 1537-2197. JSTOR 2657015. PMID 11222247.
- ^ Триверс, Роберт (1988). «Половые различия в скорости рекомбинации и полового отбора». В Michod, Richard E .; Левин, Брюс Р. (ред.). Эволюция секса. Сандерленд, Массачусетс: Sinauer Associates Inc., стр.279. ISBN 978-0-87893-459-1.
- ^ Докинз, Ричард (1988). «Взрывы и спирали». В: Слепой часовщик. Лондон: Penguin Books. С. 195–220. ISBN 978-0-14-014481-9.
- ^ Ланде, Р. (1981). «Модели видообразования путем полового отбора по полигенным признакам». Труды Национальной академии наук США. 78 (6): 3721–3725. Bibcode:1981PNAS ... 78.3721L. Дои:10.1073 / pnas.78.6.3721. ЧВК 319643. PMID 16593036.