Естественный отбор - Natural selection


Естественный отбор это дифференциальная выживаемость и воспроизводство особей из-за различий в фенотип. Это ключевой механизм эволюция, изменение наследственный черты характеристика численность населения через поколения. Чарльз Дарвин популяризировал термин "естественный отбор", противопоставляя его искусственный отбор, что, по его мнению, является преднамеренным, тогда как естественный отбор - нет.
Вариация существует во всех популяциях организмы. Отчасти это происходит потому, что случайный мутации возникают в геном отдельного организма, и их потомство могут унаследовать такие мутации. На протяжении всей жизни людей их геномы взаимодействуют с окружающей средой, вызывая изменения в чертах. Окружение генома включает молекулярную биологию в клетка, другие клетки, другие люди, популяции, разновидность, а также абиотическая среда. Поскольку особи с определенными вариантами признака, как правило, выживают и размножаются больше, чем люди с другими менее успешными вариантами, популяция эволюционирует. Другие факторы, влияющие на репродуктивный успех, включают: половой отбор (теперь часто включается в естественный отбор) и отбор плодовитости.
Естественный отбор воздействует на фенотип, характеристики организма, которые действительно взаимодействуют с окружающей средой, но генетический (наследственная) основа любого фенотипа, придающая этому фенотипу репродуктивное преимущество, может стать чаще встречается в популяции. Со временем в результате этого процесса могут появиться группы, специализирующиеся на определенных экологические ниши (микроэволюция ) и может в конечном итоге привести к видообразование (появление новых видов, макроэволюция ). Другими словами, естественный отбор - ключевой процесс в эволюции популяции.
Естественный отбор - краеугольный камень современного биология. Концепция, опубликованная Дарвином и Альфред Рассел Уоллес в совместное представление документов 1858 г., был разработан во влиятельной книге Дарвина 1859 г. О происхождении видов путем естественного отбора или о сохранении избранных рас в борьбе за жизнь. Он описал естественный отбор как аналог искусственного отбора - процесса, с помощью которого животные и растения с признаками, которые селекционеры считают желательными, систематически отдают предпочтение воспроизводству. Концепция естественного отбора первоначально возникла в отсутствие действующей теории наследственности; На момент написания Дарвина наука еще не разработала современные теории генетики. Союз традиционных Дарвиновская эволюция с последующими открытиями в классическая генетика сформировал современный синтез середины 20 века. Добавление молекулярная генетика привело к эволюционная биология развития, который объясняет эволюцию на молекулярном уровне. Пока генотипы может медленно меняться случайным образом генетический дрейф, естественный отбор остается основным объяснением адаптивная эволюция.
Историческое развитие
Додарвиновские теории

Некоторые философы классическая эпоха, включая Эмпедокл[1] и его интеллектуальный преемник, Римский поэт Лукреций,[2] выразил идею о том, что природа произвольно производит огромное количество разнообразных существ и что выживают только те существа, которым удается обеспечивать себя и успешно воспроизводиться. Идея Эмпедокла о том, что организмы возникли исключительно в результате случайного действия таких причин, как жара и холод, подверглась критике со стороны Аристотель в Книге II Физика.[3] Он положил естественный телеология на его месте, и считал, что форма была достигнута с определенной целью, ссылаясь на закономерность наследственности видов в качестве доказательства.[4][5] Тем не менее он принял в его биологии что новые виды животных, чудовища (τερας), может встречаться в очень редких случаях (Генерация животных, Книга IV).[6] Как указано в издании Дарвина 1872 г. Происхождение видов, Аристотель размышлял, могли ли разные формы (например, зубы) возникнуть случайно, но выжили только полезные формы:
Так что же мешает различным частям [тела] иметь эту просто случайную связь в природе? поскольку зубы, например, растут по необходимости, передние острые, приспособленные для деления, а измельчители плоские и пригодные для жевания пищи; так как они были сделаны не ради этого, а в результате несчастного случая. То же самое и с другими частями, в которых, кажется, существует приспособление к концу. Следовательно, где бы все вещи вместе (то есть все части одного целого) ни происходили, как если бы они были созданы ради чего-то, они сохранялись, будучи надлежащим образом составленными внутренней спонтанностью, и все, что бы ни было таким образом образовано , погибли и все еще погибают.
— Аристотель, Физика, Книга II, Глава 8[7]
Но Аристотель отверг эту возможность в следующем абзаце, пояснив, что он говорит о развитие животных как эмбрионов с фразой «либо всегда, либо обычно», а не с происхождением вида:
... Однако невозможно, чтобы это было истинным мнением. Ведь зубы и все другие естественные вещи либо неизменно, либо обычно возникают определенным образом; но не об одном из результатов случайности или спонтанности. Мы не приписываем случайности или простому совпадению частоту дождя зимой, но частый дождь летом мы делаем; ни тепла в собачьи дни, а только зимой. Если тогда согласовано, что вещи являются результатом совпадения или конечного результата, а они не могут быть результатом совпадения или спонтанности, из этого следует, что они должны быть результатом; и что все такие вещи происходят от природы, даже сторонники теории, которая находится перед нами, согласятся. Следовательно, действие для достижения цели присутствует в вещах, которые возникают и существуют по своей природе.
— Аристотель, Физика, Книга II, Глава 8[8]
В борьба за существование позже был описан Исламский писатель Аль-Джахиз в 9 веке.[9][10][11]
Классические аргументы были вновь введены в 18 веке Пьер Луи Мопертюи[12] и другие, в том числе дед Дарвина, Эразм Дарвин.
До начала 19 века преобладающее мнение в Западные общества заключалась в том, что различия между особями вида были неинтересными отклонениями от их Платонические идеалы (или же тип ) из созданные виды. Однако теория униформизм в геологии продвигали идею о том, что простые слабые силы могут действовать непрерывно в течение длительных периодов времени, вызывая радикальные изменения в земной шар пейзаж. Успех этой теории повысил осведомленность об огромных масштабах геологическое время и сделал правдоподобным идею о том, что крошечные, практически незаметные изменения в последовательных поколениях могут иметь последствия в масштабе различий между видами.[13]
Зоолог начала XIX века Жан-Батист Ламарк предложил наследование приобретенных характеристик как механизм эволюционного изменения; адаптивные черты, приобретенные организмом в течение его жизни, могут быть унаследованы потомством этого организма, в конечном итоге вызывая трансмутация видов.[14] Эта теория, Ламаркизм, оказал влияние на советского биолога Трофим Лысенко антагонизм господствующей генетической теории еще в середине 20 века.[15]
Между 1835 и 1837 годами зоолог Эдвард Блит работали над областью вариации, искусственного отбора и того, как подобный процесс происходит в природе. Дарвин признал идеи Блайта в первой главе об изменении О происхождении видов.[16]
Теория Дарвина
В 1859 году Чарльз Дарвин изложил свою теорию эволюции путем естественного отбора как объяснение приспособление и видообразование. Он определил естественный отбор как «принцип, по которому каждое небольшое изменение [признака], если оно полезно, сохраняется».[17] Идея была простой, но действенной: люди, лучше всего приспособленные к окружающей среде, с большей вероятностью выживут и будут воспроизводиться. Пока между ними есть некоторые различия, и эти различия наследственный неизбежен отбор особей с наиболее выгодными вариациями. Если вариации наследуются, то дифференциальный репродуктивный успех ведет к прогрессивной эволюции конкретных население вида, а популяции, которые эволюционируют и становятся достаточно разными, в конечном итоге становятся разными видами.[18][19]

Идеи Дарвина были вдохновлены наблюдениями, которые он сделал на второй рейс HMS Бигль (1831–1836), а также работой политического экономиста, Томас Роберт Мальтус, кто в Очерк принципа народонаселения (1798 г.), отметил, что население (если не отмечено) увеличивается экспоненциально, тогда как запасы пищи растут только арифметически; таким образом, неизбежные ограничения ресурсов будут иметь демографические последствия, ведущие к «борьбе за существование».[20] Когда Дарвин прочитал Мальтуса в 1838 году, его работа уже была увлечена его работой. натуралист ценить «борьбу за существование» в природе. Ему пришло в голову, что по мере того, как население перерастет ресурсы, «благоприятные вариации будут иметь тенденцию сохраняться, а неблагоприятные - уничтожаться. Результатом этого будет образование новых видов».[21] Дарвин писал:
Если на протяжении долгих веков и при различных условиях жизни органические существа вообще различаются в нескольких частях своей организации, и я думаю, это не может быть оспорено; если из-за высоких геометрических возможностей увеличения каждого вида в каком-то возрасте, сезоне или году будет жестокая борьба за жизнь, и это, конечно, не может быть оспорено; Затем, учитывая бесконечную сложность отношений всех органических существ друг к другу и к условиям их существования, что приводит к тому, что бесконечное разнообразие в структуре, строении и привычках приносит им пользу, я думаю, что это было бы весьма необычным фактом. если бы никогда не возникало никаких вариаций, полезных для собственного благополучия каждого существа, точно так же, как многие вариации были полезны для человека. Но если изменения, полезные для какого-либо органического существа, действительно происходят, несомненно, у индивидуумов, охарактеризованных таким образом, будут наилучшие шансы на сохранение в борьбе за жизнь; и из-за строгого принципа наследования они будут иметь тенденцию производить потомство с аналогичными характеристиками. Этот принцип сохранения я назвал для краткости естественным отбором.
— Дарвин резюмирует естественный отбор в четвертой главе книги. О происхождении видов[22]
Получив свою теорию, Дарвин тщательно собирал и уточнял доказательства, прежде чем обнародовать свою идею. Он был в процессе написания своей «большой книги», чтобы представить свои исследования, когда натуралист Альфред Рассел Уоллес независимо задумал принцип и описал его в эссе, которое он послал Дарвину, чтобы переслать его Чарльз Лайель. Лайель и Джозеф Далтон Хукер решил представить свое эссе вместе с неопубликованными работами, которые Дарвин послал коллегам-натуралистам, и О склонности видов к формированию разновидностей; и о сохранении разновидностей и видов естественными средствами отбора был зачитан Линнеевское общество Лондона объявляя о совместном открытии принципа в июле 1858 г.[23] Дарвин опубликовал подробный отчет о своих доказательствах и выводах в О происхождении видов в 1859 г. В 3-м издании 1861 г. Дарвин признал, что другие, например Уильям Чарльз Уэллс в 1813 г. и Патрик Мэтью в 1831 г. - предложил аналогичные идеи, но не развил их и не представил в заметных научных публикациях.[24]

Дарвин думал о естественном отборе по аналогии с тем, как фермеры отбирают сельскохозяйственные культуры или домашний скот для разведения, что он назвал "искусственный отбор "; в своих ранних рукописях он ссылался на" Природу ", которая будет осуществлять отбор. В то время другие механизмы эволюции, такие как эволюция путем генетического дрейфа, еще не были четко сформулированы, и Дарвин полагал, что отбор, вероятно, был лишь частью рассказ: «Я убежден, что естественный отбор был основным, но не исключительным средством модификации».[25] В письме Чарльзу Лайелю в сентябре 1860 года Дарвин сожалел об использовании термина «естественный отбор», предпочитая термин «естественное сохранение».[26]
Для Дарвина и его современников естественный отбор был по сути синонимом эволюции путем естественного отбора. После публикации О происхождении видов,[27] образованные люди считали, что эволюция в той или иной форме происходила. Однако естественный отбор как механизм оставался спорным, отчасти потому, что он воспринимался как слишком слабый, чтобы объяснить диапазон наблюдаемых характеристик живых организмов, а отчасти потому, что даже сторонники эволюции возражали против его «неуправляемости» и неуправляемости.прогрессивный природа,[28] ответ, который был охарактеризован как наиболее значимое препятствие для принятия идеи.[29] Однако некоторые мыслители с энтузиазмом восприняли естественный отбор; прочитав Дарвина, Герберт Спенсер ввел фразу выживание сильнейшего, который стал популярным изложением теории.[30][31] Пятое издание О происхождении видов опубликованная в 1869 году, включала в себя фразу Спенсера как альтернативу естественному отбору с указанием похвалы: «Но выражение, часто используемое г-ном Гербертом Спенсером из книги« Выживание наиболее приспособленных », является более точным, а иногда и столь же удобным».[32] Хотя эту фразу до сих пор часто используют небиологи, современные биологи избегают ее, потому что она тавтологический если «наиболее приспособленный» означает «функционально превосходящий» и применяется к отдельным лицам, а не рассматривается как усредненное количество по популяциям.[33]
Современный синтез
Естественный отбор в решающей степени опирается на идею наследственности, но он был разработан раньше, чем основные концепции генетика. Хотя Моравский монах Грегор Мендель, отец современной генетики, был современником Дарвина, его работы оставались неизвестными и были вновь обнаружены только в 1900 году.[34] В начале 20 века интеграция эволюции с Законы Менделя наследования, так называемые современный синтез, ученые обычно соглашались с естественным отбором.[35][36] Синтез вырос из достижений в различных областях. Рональд Фишер разработал необходимый математический язык и написал Генетическая теория естественного отбора (1930).[37] Дж. Б. С. Холдейн ввел понятие «издержки» естественного отбора.[38][39]Сьюэлл Райт выяснил природу отбора и адаптации.[40]В его книге Генетика и происхождение видов (1937), Феодосий Добжанский установил идею, что мутация, когда-то рассматривался как соперник к отбору, фактически предоставившим сырье для естественного отбора путем создания генетического разнообразия.[41][42]
Второй синтез

Эрнст Майр признал ключевое значение репродуктивная изоляция для видообразования в его Систематика и происхождение видов (1942).[44]В. Д. Гамильтон задумал родственный отбор в 1964 г.[45][46] Этот синтез закрепил естественный отбор как основу эволюционной теории, которой он остается и сегодня. Второй синтез был осуществлен в конце 20-го века благодаря достижениям в молекулярная генетика, создавая поле эволюционная биология развития ("эво-дево"), который пытается объяснить эволюцию форма с точки зрения программы генетического регулирования которые контролируют развитие эмбриона на молекулярном уровне. Под естественным отбором здесь понимается воздействие на эмбриональное развитие, изменяющее морфологию взрослого тела.[47][48][49][50]
Терминология
Период, термин естественный отбор чаще всего определяется как оперирование наследственными признаками, поскольку они непосредственно участвуют в эволюции. Однако естественный отбор «слепой» в том смысле, что изменения фенотипа могут дать репродуктивное преимущество независимо от того, наследуется ли признак. Следуя первичному использованию Дарвина, этот термин используется для обозначения как эволюционных последствий слепого отбора, так и его механизмов.[27][37][51][52] Иногда бывает полезно четко различать механизмы отбора и его эффекты; когда это различие важно, ученые определяют «(фенотипический) естественный отбор» конкретно как «те механизмы, которые способствуют отбору воспроизводящихся особей», независимо от того, является ли основа отбора наследственной.[53][54][55] Признаки, которые приводят к большему репродуктивному успеху организма, называются выбран для, а те, которые снижают успех, выбран против.[56]
Механизм
Наследственная вариация, дифференциальное воспроизводство

Естественная изменчивость происходит среди особей любой популяции организмов. Некоторые различия могут улучшить шансы человека на выживание и воспроизводство, так что скорость его воспроизводства в течение всей жизни увеличивается, что означает, что он оставляет больше потомства. Если черты, которые дают этим людям репродуктивное преимущество, также наследственный, то есть передаются от родителей к потомству, тогда будет дифференциальное воспроизводство, то есть немного более высокая доля быстрых кроликов или эффективных водорослей в следующем поколении. Даже если репродуктивное преимущество очень невелико, через многие поколения в популяции становится доминирующим любой выгодный наследственный признак. Таким образом окружающая среда организма «отбирает» черты, дающие репродуктивное преимущество, вызывая эволюционные изменения, как описал Дарвин.[57] Это создает видимость цели, но при естественном отборе преднамеренного выбора нет.[а] Искусственный отбор - это целенаправленный где естественного отбора нет, хотя биологи часто используют телеологический язык описать это.[58]
В моль перечная существует как в светлых, так и в темных тонах в Великобритании, но в Индустриальная революция, многие деревья, на которых покоилась моль, почернели от сажа, давая темным мотылькам преимущество в укрытии от хищников. Это дало темным мотылькам больше шансов выжить и дать потомство темного окраса, и всего за пятьдесят лет с момента поимки первого темного мотылька почти все бабочки промышленного Манчестер были темными. Баланс был полностью изменен эффектом Закон о чистом воздухе 1956 года, и темные бабочки снова стали редкими, демонстрируя влияние естественного отбора на эволюция пяденицы.[59] Недавнее исследование с использованием анализа изображений и моделей птичьего зрения показывает, что бледные люди более точно соответствуют фону лишайников, чем темные морфы, и впервые количественно оценивают камуфляж моли хищничество риск.[60]
Фитнес
Концепция приспособленности занимает центральное место в естественном отборе. В широком смысле, люди, которые более «подходят», имеют больший потенциал для выживания, как в известной фразе «выживание сильнейшего ", но точное значение этого термина гораздо более тонкое. Современная эволюционная теория определяет приспособленность не по продолжительности жизни организма, а по тому, насколько успешно он размножается. Если организм живет вдвое дольше, чем другие представители его вида, но имеет вдвое больше потомков, доживающих до взрослого возраста, его гены становятся более распространенными среди взрослого населения следующего поколения. Хотя естественный отбор действует на особей, эффекты случайности означают, что приспособленность действительно может быть определена только «в среднем» для особей внутри Приспособленность к определенному генотипу соответствует среднему эффекту для всех лиц с этим генотипом.[61]Необходимо проводить различие между концепцией «выживание наиболее приспособленных» и «улучшение физической формы». «Выживание наиболее приспособленных» не дает «улучшения приспособленности», а лишь представляет собой удаление менее приспособленных вариантов из популяции. Математический пример «выживания наиболее приспособленных» дан Холдейном в его статье «Цена естественного отбора».[62] Холдейн называл этот процесс «замещением» или, чаще в биологии, «фиксацией». Это правильно описывается дифференциальной выживаемостью и размножением особей из-за различий в фенотипе. С другой стороны, «улучшение приспособленности» не зависит от дифференциальной выживаемости и воспроизводства особей из-за различий в фенотипах, оно зависит от абсолютной выживаемости конкретного варианта. Вероятность того, что полезная мутация произойдет у какого-либо члена популяции, зависит от общего количества репликаций этого варианта. Математику «улучшения фитнеса» описал Клейнман. [63] Эмпирический пример «улучшения физической формы» дает эксперимент Кишони Мега-пластина. [64] В этом эксперименте «улучшение приспособленности» зависит от количества повторений конкретного варианта для появления нового варианта, способного расти в области следующей более высокой концентрации лекарственного средства. Для этого «улучшения физической формы» фиксации или замены не требуется. С другой стороны, «улучшение физической формы» может происходить в среде, где также действует «выживание наиболее приспособленных». Ричард Ленски классика Кишечная палочка долгосрочный эволюционный эксперимент является примером адаптации в конкурентной среде («улучшение физической формы» во время «выживания наиболее приспособленных»).[65] Вероятность полезной мутации, происходящей на каком-либо члене линии, чтобы улучшить приспособленность, снижается из-за конкуренции. Вариант, который является кандидатом на полезную мутацию в этой среде с ограниченной несущей способностью, должен сначала превзойти «менее подходящие» варианты, чтобы накопить необходимое количество репликаций, чтобы существовала разумная вероятность возникновения этой полезной мутации.[66]
Конкуренция
В биологии конкуренция - это взаимодействие между организмами, при котором приспособленность одного снижается из-за присутствия другого. Это может быть потому, что оба полагаются на ограничено поставка ресурса, такого как еда, вода или территория.[67] Конкуренция может быть в или же между видами, и может быть прямым или косвенным.[68] Виды, менее подходящие для соревнований, должны теоретически либо адаптироваться, либо вымереть, поскольку конкуренция играет важную роль в естественном отборе, но, согласно теории «места для передвижения», она может быть менее важной, чем расширение среди более крупных клады.[68][69]
Конкурс моделируется r / K теория отбора, который основан на Роберт Макартур и Э. О. Уилсон работает над островная биогеография.[70] Согласно этой теории, давление отбора ведет эволюцию в одном из двух стереотипных направлений: р- или же K-выбор.[71] Эти условия, р и K, можно проиллюстрировать в виде логистическая модель из динамика населения:[72]

куда р это скорость роста населения (N), и K это грузоподъемность его местной окружающей среды. Обычно р-выбранные виды эксплуатируют пустые ниши, и дают много потомства, каждое с относительно низким вероятность дожить до взрослой жизни. В отличие, K- отдельные виды являются сильными конкурентами в переполненных нишах, и вкладывать деньги в большей степени у гораздо меньшего числа потомков, каждое из которых имеет относительно высокую вероятность дожить до взрослого возраста.[72]
Классификация

2, стабилизирующий отбор: средний уровень предпочтительнее крайностей.
3: разрушительный отбор: крайности предпочтительнее промежуточных.
Ось X: фенотипический признак
Ось Y: количество организмов
Группа A: исходное население
Группа B: после отбора
Естественный отбор может воздействовать на любые наследуемые фенотипический признак,[73] а давление отбора может быть вызвано любым аспектом окружающей среды, включая половой отбор и конкуренция с представителями того же или другого вида.[74][75] Однако это не означает, что естественный отбор всегда направлен и приводит к адаптивной эволюции; естественный отбор часто приводит к поддержанию статус-кво за счет исключения менее подходящих вариантов.[57]
Отбор можно классифицировать несколькими различными способами, например, по его влиянию на признак, на генетическое разнообразие, по стадии жизненного цикла, на которой он действует, по единице отбора или по ресурсу, за который идет конкуренция.
По влиянию на черту
Отбор по-разному влияет на черты характера. Стабилизирующий отбор действует, чтобы удерживать признак на стабильном оптимуме, и в простейшем случае все отклонения от этого оптимума выборочно невыгодны. Направленный выбор выступает за крайние значения черты характера. Необычный подрывной отбор также действует в переходные периоды, когда текущий режим неоптимален, но изменяет признак в более чем одном направлении. В частности, если признак количественный и одномерный тогда предпочтение отдается как более высокому, так и низкому уровню черт. Подрывной отбор может предшествовать видообразование.[57]
По влиянию на генетическое разнообразие
В качестве альтернативы выделение можно разделить в зависимости от его влияния на генетическое разнообразие. Очищающий или отрицательный отбор действует по удалению генетической изменчивости из популяции (и этому противостоит de novo мутация, который вводит новую вариацию.[76][77] В отличие, балансирующий выбор действует для поддержания генетической изменчивости в популяции, даже при отсутствии de novo мутация, отрицательная частотно-зависимый выбор. Один из механизмов для этого: преимущество гетерозиготы, где люди с двумя разными аллелями имеют избирательное преимущество перед людьми с одним аллелем. Полиморфизм у человека Группа крови АВО locus был объяснен таким образом.[78]
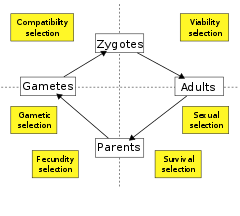
По стадии жизненного цикла
Другой вариант - классифицировать выбор по жизненный цикл этап, на котором он действует. Некоторые биологи выделяют всего два типа: отбор по жизнеспособности (или выживаемости), который действует для увеличения вероятности выживания организма, и отбора плодовитости (или фертильности, или репродуктивного), который действует для увеличения скорости воспроизводства при условии выживаемости. Другие разделяют жизненный цикл на дополнительные составляющие отбора. Таким образом, отбор на жизнеспособность и выживаемость может быть определен отдельно и, соответственно, как действие, направленное на повышение вероятности выживания до и после достижения репродуктивного возраста, в то время как отбор по плодовитости можно разделить на дополнительные подкомпоненты, включая половой отбор, гаметический отбор, воздействие на гамета выживание и выбор совместимости, действующие на зигота формирование.[79]
По единице выбора
Выбор также можно классифицировать по уровню или единица выбора. Индивидуальный отбор действует на индивида в том смысле, что адаптации служат «на благо» индивида и являются результатом отбора среди индивидов. Выбор гена действует непосредственно на уровне гена. В родственный отбор и внутригеномный конфликт, отбор на уровне генов дает более подходящее объяснение лежащего в основе процесса. Выбор группы если это происходит, действует на группы организмов, исходя из предположения, что группы реплицируются и мутируют аналогично генам и индивидуумам. Продолжаются дискуссии о том, в какой степени групповой отбор происходит в природе.[80]
По ресурсам, за которые идет конкуренция

Наконец, выбор можно классифицировать по ресурс конкурируют за. Половой отбор является результатом конкуренции за партнеров. Половой отбор обычно осуществляется путем отбора плодовитости, иногда в ущерб жизнеспособности. Экологический отбор естественный отбор с помощью любых средств, кроме полового, таких как родственный отбор, конкуренция и детоубийство. Вслед за Дарвином естественный отбор иногда определяется как экологический отбор, и в этом случае половой отбор считается отдельным механизмом.[83]
Половой отбор, впервые сформулированный Дарвином (на примере павлин хвост)[81] относится конкретно к соревнованию за товарищей,[84] который может быть внутрисексуальный, между людьми одного пола, то есть конкуренция между мужчинами, или интерсексуальный, где один пол выбирает товарищей, чаще всего с показом самцов и выбором самок.[85] Однако у некоторых видов выбор партнера в первую очередь осуществляется самцами, как у некоторых рыб этого семейства. Syngnathidae.[86][87]
Фенотипические признаки могут быть отображается у одного пола и желанны у другого, вызывая положительный отзыв петля называется Беглый рыбак например, экстравагантное оперение некоторых самцов птиц, таких как павлин.[82] Альтернативная теория, предложенная тем же Рональд Фишер в 1930 г. гипотеза сексуального сына, что матери хотят иметь беспорядочных сыновей, чтобы дать им большое количество внуков, и поэтому выбирают беспорядочных отцов для своих детей. Агрессия между представителями одного пола иногда ассоциируется с очень отличительными чертами, такими как рога олени, которые используются в бою с другими оленями. В более общем плане интрасексуальный отбор часто ассоциируется с половой диморфизм, включая различия в размерах тела между самцами и самками одного вида.[85]
Гонки вооружений
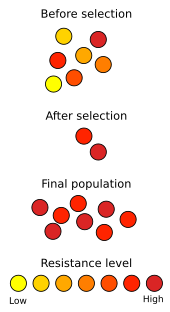
Естественный отбор проявляется в действии в развитии устойчивость к антибиотикам в микроорганизмы. С момента открытия пенициллин в 1928 г., антибиотики использовались для борьбы с бактериальными заболеваниями. Широко распространенное злоупотребление антибиотиками привело к отбору микробной устойчивости к антибиотикам в клинических условиях до такой степени, что метициллин-устойчивый Золотистый стафилококк (MRSA) был описан как «супербактерия» из-за угрозы, которую он представляет для здоровья, и его относительной неуязвимости для существующих лекарств.[88] Стратегии реагирования обычно включают использование различных, более сильных антибиотиков; однако новые напряжения MRSA, устойчивые даже к этим препаратам.[89] Это эволюционная гонка вооружений, в котором бактерии вырабатывают штаммы, менее чувствительные к антибиотикам, в то время как медицинские исследователи пытаются разработать новые антибиотики, которые могут их убить. Аналогичная ситуация возникает с устойчивость к пестицидам у растений и насекомых. Гонка вооружений не обязательно вызвана человеком; хорошо задокументированный пример включает распространение гена у бабочки Hypolimnas bolina подавление деятельности по убийству мужчин путем Вольбахия бактерии-паразиты на острове Самоа, где распространение гена, как известно, произошло всего за пять лет[90][91]
Эволюция посредством естественного отбора
Предпосылкой для естественного отбора, приводящего к адаптивной эволюции, новым признакам и видообразованию, является наличие наследственной генетической изменчивости, которая приводит к различиям в приспособленности. Генетическая изменчивость - результат мутаций, генетические рекомбинации и изменения в кариотип (количество, форма, размер и внутреннее расположение хромосомы ). Любое из этих изменений может иметь очень выгодный или крайне невыгодный эффект, но большие эффекты случаются редко. В прошлом большинство изменений в генетическом материале считалось нейтральным или близким к нейтральному, потому что они происходили в некодирующая ДНК или привело к синонимичная замена. Однако многие мутации в некодирующей ДНК имеют пагубные последствия.[92][93] Хотя как частота мутаций, так и среднее влияние мутаций на приспособляемость зависят от организма, большинство мутаций у людей немного вредны.[94]
Некоторые мутации происходят в «инструментарий» или регуляторные гены. Их изменения часто имеют большое влияние на фенотип человека, потому что они регулируют функцию многих других генов. Большинство (но не все) мутаций в регуляторных генах приводят к нежизнеспособным эмбрионам. Некоторые нелетальные регуляторные мутации возникают в Гены HOX у людей, что может привести к шейное ребро[95] или же полидактилия, увеличение количества пальцев рук или ног.[96] Когда такие мутации приводят к более высокой приспособленности, естественный отбор благоприятствует этим фенотипам, и новый признак распространяется в популяции. Установленные признаки не являются неизменными; черты, обладающие высокой пригодностью в одном контексте окружающей среды, могут оказаться гораздо менее подходящими при изменении условий окружающей среды. В отсутствие естественного отбора, направленного на сохранение такой черты, она становится более изменчивой и со временем ухудшается, что может привести к рудиментарный проявление признака, также называемого эволюционный багаж. Во многих случаях внешне рудиментарная структура может сохранять ограниченную функциональность или может быть использована для получения других полезных свойств в явлении, известном как преадаптация. Известный пример рудиментарной структуры, глаз слепой землекоп, как полагают, сохраняет функцию в фотопериод восприятие.[97]
Видообразование
Для видообразования требуется степень репродуктивная изоляция - то есть уменьшение потока генов. Однако это неотъемлемая часть концепции разновидность который гибриды отбираются против эволюции репродуктивной изоляции - проблемы, признанной Дарвином. Проблема не возникает в аллопатрический видообразование с географически разделенными популяциями, которые могут расходиться с различными наборами мутаций. Э. Б. Поултон В 1903 году осознал, что репродуктивная изоляция может развиваться через дивергенцию, если каждая линия приобретает разные несовместимые аллели одного и того же гена. Тогда отбор против гетерозиготы непосредственно приведет к репродуктивной изоляции, что приведет к Модель Бейтсона – Добжанского – Мюллера., далее разработанная Х. Аллен Орр[98] и Сергей Гаврилец.[99] С подкрепление тем не менее, естественный отбор может способствовать увеличению презиготической изоляции, напрямую влияя на процесс видообразования.[100]
Генетическая основа
Генотип и фенотип
Естественный отбор влияет на фенотип или физические характеристики организма. Фенотип определяется генетическим составом (генотипом) организма и окружающей средой, в которой он живет. Когда разные организмы в популяции обладают разными версиями гена определенного признака, каждая из этих версий известна как аллель. Именно эта генетическая изменчивость лежит в основе различий в фенотипах. Примером может служить ABO группа крови антигены у людей, где фенотипом управляют три аллеля.[101]
Некоторые черты регулируются только одним геном, но на большинство черт влияют взаимодействия многих генов. Вариация одного из многих генов, которые вносят вклад в признак, может иметь лишь небольшое влияние на фенотип; вместе эти гены могут производить континуум возможных фенотипических значений.[102]
Направленность отбора
Когда некоторый компонент признака является наследуемым, отбор изменяет частоты различных аллелей или вариантов гена, который производит варианты признака. Отбор можно разделить на три класса в зависимости от его влияния на частоты аллелей: направленный, стабилизирующий, и подрывной отбор.[103] Направленный отбор происходит, когда аллель имеет большую приспособленность, чем другие, так что он увеличивается по частоте, получая все большую долю в популяции. Этот процесс может продолжаться до тех пор, пока аллель не станет фиксированный и все население разделяет фенотип приспособленности.[104] Гораздо более распространенным является стабилизирующий отбор, который снижает частоту аллелей, которые оказывают пагубное влияние на фенотип, то есть производят организмы с более низкой приспособленностью. Этот процесс может продолжаться до тех пор, пока аллель не будет исключен из популяции. Стабилизирующий отбор сохраняет функциональные генетические особенности, такие как гены, кодирующие белок или же регуляторные последовательности со временем путем выборочного давления на вредоносные варианты.[105] Подрывной (или диверсифицирующий) отбор - это отбор, в котором предпочтение отдается экстремальным значениям признаков над промежуточными значениями признаков. Подрывной отбор может вызвать симпатрическое видообразование через перегородка ниши.
Некоторые формы балансирующий выбор не приводят к фиксации, но поддерживают аллель на промежуточных частотах в популяции. Это может произойти в диплоид виды (с парами хромосом), когда гетерозиготный особи (только с одной копией аллеля) имеют более высокую приспособленность, чем гомозиготные особи (с двумя копиями). Это называется гетерозиготным преимуществом или чрезмерным преобладанием, наиболее известным примером которого является устойчивость к малярии у людей, гетерозиготных по серповидно-клеточная анемия. Поддержание аллельной изменчивости также может происходить за счет подрывной или диверсифицирующий отбор, что благоприятствует генотипам, отклоняющимся от среднего в любом направлении (то есть в противоположность чрезмерному преобладанию), и может привести к бимодальное распределение ценностей черт. Наконец, уравновешивающий отбор может происходить посредством частотно-зависимого отбора, когда приспособленность одного конкретного фенотипа зависит от распределения других фенотипов в популяции. Принципы теория игры были применены для понимания распределений приспособленности в этих ситуациях, особенно при изучении родственного отбора и эволюции взаимный альтруизм.[106][107]
Отбор, генетическая изменчивость и дрейф
Часть всех генетических вариаций функционально нейтральна, не вызывая фенотипического эффекта или значительной разницы в приспособленности. Мотоо Кимура с нейтральная теория молекулярной эволюции к генетический дрейф предполагает, что эта вариация составляет большую часть наблюдаемого генетического разнообразия.[108] Нейтральные события могут радикально уменьшить генетическую изменчивость за счет узкие места населения.[109] что, среди прочего, может вызвать эффект основателя в изначально небольших новых популяциях.[110] Когда генетическая изменчивость не приводит к различиям в приспособленности, отбор не может напрямую влиять на частоту такой вариации. В результате генетическая изменчивость на этих участках выше, чем на участках, где вариации действительно влияют на приспособленность.[103] Однако после периода отсутствия новых мутаций генетическая изменчивость на этих участках устраняется из-за генетического дрейфа. Естественный отбор уменьшает генетическую изменчивость, устраняя дезадаптированных особей и, следовательно, мутации, вызвавшие дезадаптацию. В то же время происходят новые мутации, в результате чего баланс мутации и отбора. Точный результат этих двух процессов зависит как от скорости появления новых мутаций, так и от силы естественного отбора, который является функцией того, насколько неблагоприятной окажется мутация.[111]
Генетическая связь происходит, когда места двух аллелей находятся в непосредственной близости на хромосоме. Во время образования гамет рекомбинация перетасовывает аллели. Вероятность того, что такая перестановка произойдет между двумя аллелями, обратно пропорциональна расстоянию между ними. Выборочные развертки возникают, когда аллель становится более распространенным в популяции в результате положительного отбора. По мере увеличения распространенности одного аллеля тесно связанные аллели также могут становиться более распространенными за счет "генетический автостоп ", независимо от того, являются ли они нейтральными или даже слегка вредными. Сильное избирательное сканирование приводит к области генома, в которой гаплотип (аллель и его соседи), по сути, единственные, которые существуют в популяции. Выборочные развертки могут быть обнаружены путем измерения нарушение равновесия по сцеплению, или данный гаплотип чрезмерно представлен в популяции. Так как избирательное сканирование также приводит к отбору соседних аллелей, наличие блока сильного неравновесия по сцеплению может указывать на «недавнее» избирательное сканирование около центра блока.[112]
Выбор фона является противоположностью выборочной развертки. Если конкретный сайт подвергается сильному и стойкому очищающему отбору, связанные вариации, как правило, отсеиваются вместе с ним, создавая в геноме область с низкой общей вариабельностью. Поскольку фоновый отбор является результатом новых вредных мутаций, которые могут случайным образом происходить в любом гаплотипе, он не создает четких блоков неравновесия по сцеплению, хотя при низкой рекомбинации он все же может приводить к слегка отрицательному неравновесию по сцеплению в целом.[113]
Влияние
Идеи Дарвина, наряду с идеями Адам Смит и Карл Маркс, оказал глубокое влияние на мысль XIX века, включая его радикальное заявление о том, что «тщательно сконструированные формы, столь разные друг от друга и столь сложным образом зависимые друг от друга» произошли от простейших форм жизни на основе нескольких простых принципов.[114] Это вдохновило некоторых из самых горячих сторонников Дарвина и вызвало сильнейшее сопротивление. Естественный отбор имел силу, согласно Стивен Джей Гулд, чтобы «свергнуть некоторые из самых глубоких и самых традиционных удобств западной мысли», например, веру в то, что люди занимают особое место в мире.[115]
По словам философа Дэниел Деннетт «Опасная идея Дарвина» об эволюции путем естественного отбора - это «универсальная кислота», которую нельзя ограничивать каким-либо сосудом или контейнером, поскольку она вскоре просачивается наружу, прокладывая себе путь во все более обширное окружение.[116] Таким образом, в последние десятилетия концепция естественного отбора распространилась из эволюционная биология к другим дисциплинам, в том числе эволюционные вычисления, квантовый дарвинизм, эволюционная экономика, эволюционная эпистемология, эволюционная психология, и космологический естественный отбор. Эта неограниченная применимость была названа универсальный дарвинизм.[117]
Происхождение жизни
Как возникла жизнь из неорганической материи, остается нерешенной проблемой в биологии. Одна известная гипотеза заключается в том, что жизнь впервые появилась в виде короткой самовоспроизводящейся РНК полимеры.[118] С этой точки зрения жизнь могла возникнуть, когда РНК цепи сначала испытали основные условия, как задумал Чарльз Дарвин, для работы естественного отбора. Этими условиями являются: наследственность, вариация типа, и конкуренция за ограниченные ресурсы. Фитнес раннего Репликатор РНК вероятно, было функцией адаптивных способностей, которые были врожденными (т.е. определялись нуклеотидная последовательность ) и доступность ресурсов.[119][120] Логически можно было бы выделить три основных адаптивных способности: (1) способность воспроизводить с умеренной точностью (что приводит как к наследуемости, так и к вариациям типа), (2) способность избегать распада и (3) способность приобретать и ресурсы процесса.[119][120] Эти возможности изначально определялись сложенными конфигурациями (включая конфигурации с рибозим активности) репликаторов РНК, которые, в свою очередь, кодировались бы в их индивидуальных нуклеотидных последовательностях.[121]
Клеточная и молекулярная биология
В 1881 году эмбриолог Вильгельм Ру опубликовано Der Kampf der Theile im Organismus (Борьба частей в организме), в котором он предположил, что развитие организма является результатом дарвиновской конкуренции между частями эмбриона, происходящей на всех уровнях, от молекул до органов.[122] В последние годы современная версия этой теории была предложена Жан-Жак Купец. Согласно этому клеточному дарвинизму, случайное изменение на молекулярном уровне порождает разнообразие типов клеток, тогда как межклеточные взаимодействия устанавливают характерный порядок в развивающемся эмбрионе.[123]
Социально-психологическая теория
Социальные последствия теории эволюции путем естественного отбора также стали источником непрекращающихся споров. Фридрих Энгельс, немец политический философ и соучредитель идеологии коммунизм, писал в 1872 году, что «Дарвин не знал, какую горькую сатиру он написал на человечество, и особенно на своих соотечественников, когда он показал, что свободная конкуренция, борьба за существование, которую экономисты отмечают как высшее историческое достижение, является нормальным явлением. состояние животное царство."[124] Герберт Спенсер и защитник евгеники Фрэнсис Гальтон Интерпретация естественного отбора как обязательно прогрессивного, ведущего к предполагаемому прогрессу в интеллекте и цивилизации, стала оправданием для колониализм, евгеника, и социальный дарвинизм. Например, в 1940 г. Конрад Лоренц в трудах, от которых он впоследствии отказался, использовал теорию как оправдание политики Нацистский государственный. Он писал: «... отбор по стойкости, героизму и социальной полезности ... должен осуществляться каким-то человеческим институтом, если человечество, в отсутствие факторов отбора, не должно быть уничтожено вырождением, вызванным одомашниванием. Расовая идея как основа нашего государства уже многое сделала в этом отношении ».[125] Другие разработали идеи о том, что человеческое общество и культура эволюционировать с помощью механизмов, аналогичных тем, которые применимы к эволюции видов.[126]
Совсем недавно работа антропологов и психологов привела к развитию социобиология а затем эволюционной психологии, области, которая пытается объяснить особенности человеческая психология с точки зрения адаптации к родовой среде. Самый яркий пример эволюционной психологии, особенно развитый в ранних работах Ноам Хомский а позже Стивен Пинкер, это гипотеза о том, что человеческий мозг адаптировался к приобретать то грамматический правила естественный язык.[127] Другие аспекты человеческого поведения и социальных структур, основанные на конкретных культурных нормах, таких как предотвращение инцеста к более широким моделям, таким как гендерные роли, была выдвинута гипотеза, что их происхождение аналогично адаптации к ранней среде, в которой эволюционировали современные люди. По аналогии с действием естественного отбора на гены концепция мемы - возникли «единицы культурной передачи» или культурные эквиваленты генов, подвергающихся отбору и рекомбинации, впервые описанные в этой форме Ричард Докинз в 1976 г.[128] и впоследствии расширен такими философами, как Дэниел Деннетт в качестве объяснения сложной культурной деятельности, в том числе человеческой сознание.[129]
Теория информации и систем
В 1922 г. Альфред Дж. Лотка предположил, что естественный отбор можно понимать как физический принцип, который можно описать в терминах использования энергии системой,[130][131] концепция, позже разработанная Ховард Т. Одум как принцип максимальной мощности в термодинамика, посредством чего эволюционные системы с избирательным преимуществом максимизируют скорость преобразования полезной энергии.[132]
Принципы естественного отбора вдохновили на создание множества вычислительных методов, таких как «мягкий» искусственная жизнь, которые имитируют избирательные процессы и могут быть очень эффективными при «адаптации» сущностей к среде, определенной указанным фитнес-функция.[133] Например, класс эвристический оптимизация алгоритмы известный как генетические алгоритмы, впервые Джон Генри Холланд в 1970-х годах и расширен Дэвид Э. Голдберг,[134] определить оптимальные решения путем моделирования воспроизводства и мутации популяции решений, определенных исходным распределение вероятностей.[135] Такие алгоритмы особенно полезны при решении задач, энергетический ландшафт очень грубая или имеет много локальных минимумов.[136]
В художественной литературе
Дарвиновская эволюция путем естественного отбора широко распространена в литературе, независимо от того, оптимистично ли она воспринимается с точки зрения того, как человечество может развиваться к совершенству, или пессимистично с точки зрения ужасных последствий взаимодействия человеческой природы и борьбы за выживание. Среди основных ответов: Сэмюэл Батлер пессимистический Эревон («нигде», пишется в основном наоборот). В 1893 г. Х. Г. Уэллс воображаемый "Человек года на миллион ", преобразованное естественным отбором в существо с огромной головой, глазами и сморщенным телом.[137]
Примечания
- ^ В половой отбор можно утверждать, что самка животного, выбирающая себе пару, намеревается получить лучшего партнера; нет никаких предположений о том, что у нее есть какое-либо намерение улучшить родословную, как заводчик животных.
Рекомендации
- ^ Эмпедокл 1898, На природе, Книга II
- ^ Лукреций 1916, О природе вещей, Книга V
- ^ Аристотель, Физика, Книга II, главы 4 и 8
- ^ Лир 1988, п.38
- ^ Генри, Девин (сентябрь 2006 г.). «Аристотель о механизме наследования». Журнал истории биологии. 39 (3): 425–455. Дои:10.1007 / s10739-005-3058-у. S2CID 85671523.
- ^ Ariew 2002
- ^ Дарвин 1872, п.xiii
- ^ Аристотель, Физика, Книга II, Глава 8
- ^ Циркле, Конвей (25 апреля 1941 г.). "Естественный отбор до" происхождения видов "'". Труды Американского философского общества. 84 (1): 71–123. JSTOR 984852.
- ^ Агаттер и Уитли, 2008 г., п. 43
- ^ Мухаммад А.С. Абдель, Халим (2013). «Взгляды додарвиновских мусульманских ученых на эволюцию» (PDF). Энциклопедия наук и религий: 74–77 - через Springer.
- ^ Мопертюи, Пьер Луи (1746). [«Вывод законов движения и равновесия из метафизического принципа» ]. Histoire de l'Académie Royale des Sciences et des Belles Lettres (На французском). Берлин: 267–294.CS1 maint: ref = harv (связь)
- ^ Боулер, Питер Дж. (2003). Эволюция: история идеи (3-е изд.). Беркли, Калифорния: Калифорнийский университет Press. стр.129–134. ISBN 978-0-520-23693-6. OCLC 43091892.
- ^ Ламарк 1809
- ^ Джоравский, Дэвид (январь 1959). «Советский марксизм и биология до Лысенко». Журнал истории идей. 20 (1): 85–104. Дои:10.2307/2707968. JSTOR 2707968.
- ^ Дарвин 1859, п.18
- ^ Дарвин 1859, п.61
- ^ Дарвин 1859, п.5
- ^ Холл, Брайан К .; Халльгримссон, Бенедикт (2008). Эволюция Стрикбергера (4-е изд.). Джонс и Бартлетт. С. 4–6. ISBN 978-0-7637-0066-9. OCLC 796450355.
- ^ Мальтус 1798
- ^ Дарвин 1958, п.120
- ^ Дарвин 1859, стр.126–127
- ^ Уоллес 1871
- ^ Дарвин 1861, п.xiii
- ^ Дарвин 1859, п.6
- ^ Дарвин, Чарльз (28 сентября 1860 г.). "Дарвин, К. Р. Лайелю, Чарльзу". Дарвинский заочный проект. Кембридж, Великобритания: Библиотека Кембриджского университета. Письмо 2931. Получено 1 августа 2015.
- ^ а б Дарвин 1859
- ^ Эйсли 1958
- ^ Кун 1996
- ^ Дарвин, Чарльз (5 июля 1866 г.). "Дарвин, К. Р. Уоллесу, А. Р." Дарвинский заочный проект. Кембридж, Великобритания: Библиотека Кембриджского университета. Письмо 5145. Получено 12 января 2010.
- ^ Штуке, Морис Э. (лето 2008 г.). «Лучшая защита конкуренции». Обзор закона Святого Иоанна. Ямайка, штат Нью-Йорк. 82 (3): 951–1036.
Это выживание наиболее приспособленных, которое я попытался выразить в механических терминах, и есть то, что г.Дарвин называл «естественный отбор или сохранение избранных рас в борьбе за жизнь».
—Герберт Спенсер, Принципы биологии (1864), т. 1. С. 444–445. - ^ Дарвин 1872, п.49.
- ^ Миллс, Сьюзен К .; Битти, Джон Х. (1979). «Интерпретация фитнеса по склонности» (PDF). Философия науки. 46 (2): 263–286. CiteSeerX 10.1.1.332.697. Дои:10.1086/288865. Архивировано из оригинал (PDF) 25 декабря 2015 г.. Получено 4 августа 2015.
- ^ Эмброуз, Майк. «Горох Менделя». Норидж, Великобритания: Отдел по ресурсам зародышевой плазмы, Центр Джона Иннеса. Архивировано из оригинал 14 июня 2016 г.. Получено 22 мая 2015.
- ^ Хаксли, Джулиан (1929–1930). "A B C генетики". Наука о жизни. 2. Лондон: Amalgamated Press. OCLC 3171056.
- ^ Национальная Академия Наук (1999). Наука и креационизм: взгляд из Национальной академии наук (2-е изд.). Вашингтон, округ Колумбия: Национальная академия прессы. ISBN 978-0-309-06406-4. OCLC 43803228.
- ^ а б Фишер 1930
- ^ Холдейн 1932
- ^ Холдейн, Дж. Б. С. (Декабрь 1957 г.). «Цена естественного отбора» (PDF). Журнал генетики. 55 (3): 511–524. Дои:10.1007 / BF02984069. S2CID 32233460.
- ^ Райт, Сьюэлл (1932). «Роли мутации, инбридинга, скрещивания и отбора в эволюции». Материалы VI Международного конгресса генетиков.. 1: 356–366.
- ^ Добжанский 1937
- ^ Добжанский 1951
- ^ Кэрролл, Шон Б.; Гренье, Дженнифер К .; Уэтерби, Скотт Д. (2005). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных - второе издание. Блэквелл Паблишинг. С. 66–67. ISBN 978-1-4051-1950-4.
- ^ Майр 1942 г.
- ^ Гамильтон, В. (1964). «Генетическая эволюция социального поведения. I». Журнал теоретической биологии. 7 (1): 1–16. Дои:10.1016/0022-5193(64)90038-4. PMID 5875341.
- ^ Гамильтон, В. (1964). «Генетическая эволюция социального поведения. II». Журнал теоретической биологии. 7 (1): 17–52. Дои:10.1016/0022-5193(64)90039-6. PMID 5875340.
- ^ Гилберт, Скотт Ф. (2003). «Морфогенез эволюционной биологии развития» (PDF). Международный журнал биологии развития. 47 (7–8): 467–477. PMID 14756322.
- ^ Gilbert, S.F .; Opitz, J.M .; Рафф, Р.А. (1996). «Ресинтез эволюционной биологии и биологии развития». Биология развития. 173 (2): 357–372. Дои:10.1006 / dbio.1996.0032. PMID 8605997.
- ^ Мюллер, Г. (2007). «Evo – DevO: расширение эволюционного синтеза». Природа Обзоры Генетика. 8 (12): 943–949. Дои:10.1038 / nrg2219. PMID 17984972. S2CID 19264907.
- ^ Кэрролл, Шон Б.; Гренье, Дженнифер К .; Уэтерби, Скотт Д. (2005). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных - второе издание. Блэквелл Паблишинг. п. 13. ISBN 978-1-4051-1950-4.
- ^ Уильямс 1966
- ^ Эндлер 1986
- ^ Холдейн 1954
- ^ Ланде, Рассел; Арнольд, Стеван Дж. (Ноябрь 1983 г.). «Измерение отбора по коррелированным персонажам». Эволюция. 37 (6): 1210–1226. Дои:10.2307/2408842. JSTOR 2408842. PMID 28556011.
- ^ Футуйма 2005
- ^ Трезвый 1993
- ^ а б c «Эволюция и естественный отбор». Университет Мичигана. 10 октября 2010 г.. Получено 9 ноября 2016.
- ^ «Телеологические понятия в биологии». Стэнфордская энциклопедия философии. 18 мая 2003 г.. Получено 28 июля 2016.
- ^ ван'т Хоф, Арьен Э .; Кампань, Паскаль; Ригден, Дэниел Дж; и другие. (Июнь 2016). «Мутация промышленного меланизма у британской пяденицы - переносимый элемент». Природа. 534 (7605): 102–105. Bibcode:2016Натура.534..102H. Дои:10.1038 / природа17951. PMID 27251284.
- ^ Уолтон, Оливия; Стивенс, Мартин (2018). «Модели зрения птиц и полевые эксперименты определяют выживаемость камуфляжа берёзовой бабочки». Биология коммуникации. 1: 118. Дои:10.1038 / с42003-018-0126-3. ЧВК 6123793. PMID 30271998.
- ^ Орр, Х. Аллен (август 2009 г.). «Фитнес и его роль в эволюционной генетике». Нат Рев Жене. 10 (8): 531–539. Дои:10.1038 / nrg2603. ЧВК 2753274. PMID 19546856.
- ^ Холдейн, Дж. Б. С. (Ноябрь 1992 г.). «Цена естественного отбора». Текущая наука. 63 (9/10): 612–625.
- ^ Клейнман, А. (2014). «Основы науки и математики случайных мутаций и естественного отбора». Статистика в медицине. 33 (29): 5074–5080. Дои:10.1002 / sim.6307. PMID 25244620.
- ^ Байм, М .; Либерман, Т. Д .; Kelsic, E.D .; Chait, R .; Gross, R .; Елин, И .; Кишоны, Р. (2016). «Пространственно-временная микробная эволюция на антибиотических ландшафтах». Наука. 353 (6304): 1147–51. Bibcode:2016Научный ... 353.1147B. Дои:10.1126 / science.aag0822. ЧВК 5534434. PMID 27609891.
- ^ Блаунт, Захари Д.; Borland, Christina Z .; Ленский, Ричард Э. (2008). "Историческая случайность и эволюция ключевой инновации в экспериментальной популяции кишечная палочка". Труды Национальной академии наук. 105 (23): 7899–906. Bibcode:2008PNAS..105.7899B. Дои:10.1073 / pnas.0803151105. JSTOR 25462703. ЧВК 2430337. PMID 18524956.
- ^ Хорошо, Б. Х .; Rouzine, I.M .; Балик, Д. Дж .; Hallatschek, O .; Десаи, М. М. (27 февраля 2012 г.). «Распределение фиксированных полезных мутаций и скорость адаптации в бесполых популяциях». Труды Национальной академии наук. 109 (13): 4950–4955. Дои:10.1073 / pnas.1119910109. ЧВК 3323973. PMID 22371564.
- ^ Бегон, Таунсенд и Харпер, 1996
- ^ а б Сахней, Сарда; Бентон, Майкл Дж.; Ферри, Пол А. (23 августа 2010 г.). «Связи между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше». Письма о биологии. 6 (4): 544–547. Дои:10.1098 / рсбл.2009.1024. ЧВК 2936204. PMID 20106856.
- ^ Jardine, Phillip E .; Янис, Кристина М .; Сахней, Сарда; Бентон, Майкл Дж. (1 декабря 2012 г.). «Песок, а не трава: совпадающие закономерности раннего происхождения гипсодонтии у копытных Великих равнин и Glires». Палеогеография, палеоклиматология, палеоэкология. 365–366: 1–10. Bibcode:2012ППП ... 365 .... 1Дж. Дои:10.1016 / j.palaeo.2012.09.001.
- ^ Макартур и Уилсон 2001
- ^ Пьянка, Эрик Р. (Ноябрь – декабрь 1970 г.). "На р- и K-Выбор ». Американский натуралист. 104 (940): 592–597. Дои:10.1086/282697. JSTOR 2459020.
- ^ а б Верхюльст, Пьер Франсуа (1838). "Notice sur la loi que la population suit dans son accroissement". Соответствие Mathématique et Physique (На французском). Брюссель, Бельгия. 10: 113–121. OCLC 490225808.
- ^ Циммер и Эмлен 2013
- ^ Миллер 2000, п. 8
- ^ Арнквист, Горан; Роу, Локк (2005). Сексуальный конфликт. Издательство Принстонского университета. С. 14–43. ISBN 978-0-691-12218-2. OCLC 937342534.
- ^ Лемей, Салеми и Вандам, 2009 г.
- ^ Лоу, Лоуренс (2008). «Отрицательный выбор». Природное образование. Кембридж, Массачусетс: Издательская группа Nature. OCLC 310450541.
- ^ Villanea, Fernando A .; Safi, Kristin N .; Буш, Иеремия В. (май 2015 г.). «Общая модель отрицательного частотно-зависимого отбора объясняет глобальные закономерности полиморфизма АВО человека». PLOS ONE. 10 (5): e0125003. Bibcode:2015PLoSO..1025003V. Дои:10.1371 / journal.pone.0125003. ЧВК 4422588. PMID 25946124.
- ^ а б Кристиансен 1984, стр. 65–79
- ^ Уэйд, Майкл Дж .; и другие. (2010). «Многоуровневый и родственный отбор в связанном мире». Природа. 463 (7283): E8 – E9. Bibcode:2010 Натур.463 .... 8 Вт. Дои:10.1038 / природа08809. ЧВК 3151728. PMID 20164866.
- ^ а б Дарвин, Чарльз (1859). О происхождении видов (1-е издание). Глава 4, стр. 88. «И это заставляет меня сказать несколько слов о том, что я называю половым отбором. Это зависит от ...» http://darwin-online.org.uk/content/frameset?viewtype=side&itemID=F373&pageseq=12
- ^ а б Гринфилд, доктор медицины; Alem, S .; Limousin, D .; Бейли, Н.В. (2014). «Дилемма фишеровского полового отбора: выбор партнера для получения косвенных выгод, несмотря на редкость и общую слабость генетической корреляции между предпочтениями и признаками». Эволюция. 68 (12): 3524–3536. Дои:10.1111 / evo.12542. PMID 25308282. S2CID 2619084.
- ^ Mayr 2006
- ^ Андерссон 1994
- ^ а б Хоскен, Дэвид Дж .; Хаус, Кларисса М. (январь 2011 г.). «Половой отбор». Текущая биология. 21 (2): R62 – R65. Дои:10.1016 / j.cub.2010.11.053. PMID 21256434. S2CID 18470445.
- ^ Eens, Марсель; Пинкстен, Рианна (5 октября 2000 г.). «Изменение роли пола у позвоночных: поведенческие и эндокринологические отчеты». Поведенческие процессы. 51 (1–3): 135–147. Дои:10.1016 / S0376-6357 (00) 00124-8. PMID 11074317. S2CID 20732874.
- ^ Барлоу, Джордж У. (март 2005 г.). «Как мы решаем, что у вида поменялась роль пола?». Ежеквартальный обзор биологии. 80 (1): 28–35. Дои:10.1086/431022. PMID 15884733.
- ^ Харви, Фиона; Карсон, Мэри; О'Кейн, Мэгги; Уэсли, Эндрю (18 июня 2015 г.). «Супербактерии MRSA, обнаруженные в свинине в супермаркетах, вызывают тревогу из-за рисков, связанных с сельским хозяйством». Хранитель.
- ^ Схито, Джан К. (март 2006 г.). «Важность развития устойчивости к антибиотикам у Золотистый стафилококк". Клиническая микробиология и инфекции. 12 (Дополнение s1): 3–8. Дои:10.1111 / j.1469-0691.2006.01343.x. PMID 16445718.
- ^ Шарлат, Сильвен; Хорнетт, Эмили А .; Фуллард, Джеймс Н .; и другие. (13 июля 2007 г.). «Необычайный поток в соотношении полов». Наука. 317 (5835): 214. Bibcode:2007Наука ... 317..214C. Дои:10.1126 / science.1143369. PMID 17626876. S2CID 45723069.
- ^ Моран, Грегори Дж .; и другие. (2006). «Метициллинрезистентные инфекции S. aureus у пациентов в отделении неотложной помощи». Медицинский журнал Новой Англии. 355 (7): 666–674. Дои:10.1056 / NEJMoa055356. PMID 16914702.
- ^ Крюков Григорий В .; Шмидт, Штеффен; Сюняев, Шамиль (1 августа 2005 г.). «Небольшой фитнес-эффект мутаций в высококонсервативных некодирующих областях». Молекулярная генетика человека. 14 (15): 2221–2229. Дои:10.1093 / hmg / ddi226. PMID 15994173.
- ^ Бежерано, Гилл; Фазан, Майкл; Макунин, Игорь; и другие. (28 мая 2004 г.). «Ультраконсервированные элементы в геноме человека» (PDF). Наука. 304 (5675): 1321–1325. Bibcode:2004Научный ... 304.1321B. CiteSeerX 10.1.1.380.9305. Дои:10.1126 / science.1098119. PMID 15131266. S2CID 2790337.
- ^ Эйр-Уокер, Адам; Вулфит, Меган; Фелпс, Тед (июнь 2006 г.). «Распределение фитнес-эффектов новых вредных аминокислотных мутаций у людей». Генетика. 173 (2): 891–900. Дои:10.1534 / генетика.106.057570. ЧВК 1526495. PMID 16547091.
- ^ Галис, Фритсон (апрель 1999 г.). «Почему почти у всех млекопитающих семь шейных позвонков? Hox гены и рак ". Журнал экспериментальной зоологии. 285 (1): 19–26. Дои:10.1002 / (SICI) 1097-010X (19990415) 285: 1 <19 :: AID-JEZ3> 3.0.CO; 2-Z. PMID 10327647.
- ^ Закани, Йожеф; Фроменталь-Рамейн, Екатерина; Варо, Ксавьер; Дубуль, Дени (9 декабря 1997 г.). «Регулирование количества и размера цифр задним Hox гены: дозозависимый механизм с потенциальными эволюционными последствиями ». Труды Национальной академии наук Соединенных Штатов Америки. 94 (25): 13695–13700. Bibcode:1997PNAS ... 9413695Z. Дои:10.1073 / пнас.94.25.13695. ЧВК 28368. PMID 9391088.
- ^ Саньял, Somes; Янсен, Гарри G .; де Грип, Виллем Дж .; Нево, Эвиатар; и другие. (Июль 1990 г.). "Глаз слепой кротовой крысы, Spalax ehrenbergi. Рудимент со скрытой функцией? ". Исследовательская офтальмология и визуализация. 31 (7): 1398–1404. PMID 2142147.
- ^ Орр, Х.А. (1996). "Добжанский, Бейтсон и генетика видообразования". Генетика. 144 (4): 1331–5. ЧВК 1207686. PMID 8978022.
- ^ Гаврилец, С. (2004), Фитнес-пейзажи и происхождение видов, Издательство Принстонского университета, ISBN 978-0-691-11983-0
- ^ Шулер, Ханнес; Худ, Глен Р .; Egan, Scott P .; Федер, Джеффри Л. (2016). «Способы и механизмы видообразования». Обзоры в области клеточной биологии и молекулярной медицины. 2 (3): 60–93.
- ^ McKusick, Victor A .; Гросс, Мэтью Б. (18 ноября 2014 г.). «АВО Гликозилтрансфераза; АВО». Онлайн-менделевское наследование в человеке. Национальная медицинская библиотека. Получено 7 ноября 2016.
- ^ Falconer & Mackay 1996 г.
- ^ а б Рис 2004, См. Особенно главы 5 и 6 для количественной обработки
- ^ Rieseberg, L.H .; Widmer, A .; Arntz, A.M .; Берк, Дж. М. (2002). «Направленный отбор - основная причина фенотипической диверсификации». PNAS. 99 (19): 12242–12245. Bibcode:2002PNAS ... 9912242R. Дои:10.1073 / пнас.192360899. ЧВК 129429. PMID 12221290.
- ^ Чарльзуорт Б., Ланде Р., Слаткин М. (1982). «Неодарвинистский комментарий к макроэволюции». Эволюция. 36 (3): 474–498. Дои:10.1111 / j.1558-5646.1982.tb05068.x. JSTOR 2408095. PMID 28568049. S2CID 27361293.
- ^ Гамильтон, Уильям Д. (Июль 1964 г.). «Генетическая эволюция социального поведения. II». Журнал теоретической биологии. 7 (1): 17–52. Дои:10.1016/0022-5193(64)90039-6. PMID 5875340.
- ^ Триверс, Роберт Л. (Март 1971 г.). «Эволюция реципрокного альтруизма». Ежеквартальный обзор биологии. 46 (1): 35–57. Дои:10.1086/406755. JSTOR 2822435. S2CID 19027999.
- ^ Кимура, Мотоо (1983). Нейтральная теория молекулярной эволюции. Издательство Кембриджского университета. ISBN 978-0-521-23109-1. OCLC 8776549.
- ^ Робинсон, Ричард, изд. (2003). "Узкое место населения". Генетика. 3. Ссылка Macmillan США. ISBN 978-0-02-865609-0. OCLC 3373856121.
- ^ Кэмпбелл, Нил А. (1996). Биология (4-е изд.). Бенджамин Каммингс. п.423. ISBN 978-0-8053-1940-8. OCLC 3138680061.
- ^ Линч, Майкл (август 2010). «Эволюция скорости мутаций». Тенденции в генетике. 26 (8): 345–352. Дои:10.1016 / j.tig.2010.05.003. ЧВК 2910838. PMID 20594608.
- ^ Смит, Джон Мейнард; Хей, Джон (1974). «Автостопный эффект благоприятного гена». Генетические исследования. 23 (1): 23–35. Дои:10.1017 / S0016672300014634. PMID 4407212.
- ^ Кейтли, Питер Д.; Отто, Сара П. (7 сентября 2006 г.). «Вмешательство среди вредных мутаций способствует сексу и рекомбинации в ограниченных популяциях». Природа. 443 (7107): 89–92. Bibcode:2006 Натур.443 ... 89K. Дои:10.1038 / природа05049. PMID 16957730. S2CID 4422532.
- ^ Дарвин 1859, п.489
- ^ Гулд, Стивен Джей (12 июня 1997 г.). «Дарвиновский фундаментализм». Нью-Йоркское обозрение книг. 44 (10).
- ^ Деннет 1995
- ^ фон Сюдов, М. (2012). От дарвиновской метафизики к пониманию эволюции эволюционных механизмов. Историко-философский анализ генного дарвинизма и универсального дарвинизма. Universitätsverlag Göttingen. ISBN 978-3-86395-006-4. OCLC 1088022023.
- ^ Эйген, Манфред; Гардинер, Уильям; Шустер, Питер; и другие. (Апрель 1981 г.). «Происхождение генетической информации». Scientific American. 244 (4): 88–92, 96, et passim. Bibcode:1981SciAm.244d..88E. Дои:10.1038 / scientificamerican0481-88. PMID 6164094.
- ^ а б Бернштейн, Харрис; Байерли, Генри К.; Хопф, Фредерик А .; и другие. (Июнь 1983 г.). «Дарвиновская динамика». Ежеквартальный обзор биологии. 58 (2): 185–207. Дои:10.1086/413216. JSTOR 2828805.
- ^ а б Michod 1999
- ^ Оргел, Лесли Э. (1987). «Эволюция генетического аппарата: обзор». Симпозиумы Колд-Спринг-Харбор по количественной биологии. 52: 9–16. Дои:10.1101 / sqb.1987.052.01.004. PMID 2456886.
- ^ Ру 1881
- ^ Купец, Жан-Жак (3 мая 2010 г.). «Клеточный дарвинизм (стохастическая экспрессия генов в дифференцировке клеток и развитии эмбриона)». Научные темы. Архивировано из оригинал 4 августа 2010 г.. Получено 11 августа 2015.
- ^ Энгельс 1964
- ^ Айзенберг, Леон (Сентябрь 2005 г.). «Какой образ у Лоренца?». Американский журнал психиатрии (Письмо редактору). 162 (9): 1760. Дои:10.1176 / appi.ajp.162.9.1760. PMID 16135651. Айзенберг цитирует перевод Durch Domestikation verursachte Störungen arteigenen Verhaltens (1940, стр. 2) Конрад Лоренц.
- ^ Уилсон 2002
- ^ Пинкер 1995
- ^ Докинз 1976, п. 192
- ^ Деннет 1991
- ^ Лотка, Альфред Дж. (Июнь 1922 г.). «Вклад в энергетику эволюции». PNAS. 8 (6): 147–151. Bibcode:1922ПНАС .... 8..147Л. Дои:10.1073 / pnas.8.6.147. ЧВК 1085052. PMID 16576642.
- ^ Лотка, Альфред Дж. (Июнь 1922 г.). «Естественный отбор как физический принцип». PNAS. 8 (6): 151–154. Bibcode:1922ПНАС .... 8..151Л. Дои:10.1073 / pnas.8.6.151. ЧВК 1085053. PMID 16576643.
- ^ Одум, Х. Т. (1995). Холл, С.А.С. (ред.). Самоорганизация и максимальное расширение возможностей. Максимальная мощность: идеи и приложения H.T. Odum. Издательство Колорадского университета.
- ^ Кауфман 1993
- ^ Гольдберг 1989
- ^ Митчелл 1996
- ^ "Генетические алгоритмы". Фармакологические науки. Получено 7 ноября 2016.
- ^ Стейблфорд, Брайан М .; Лэнгфорд, Дэвид Р. (5 июля 2018 г.). «Эволюция». Энциклопедия научной фантастики. Голланц. Получено 24 июля 2018.
Источники
- Agutter, Paul S .; Уитли, Денис Н. (2008). Размышляя о жизни: история и философия биологии и других наук. Дордрехт, Нидерланды; Лондон: Springer Science + Business Media. ISBN 978-1-4020-8865-0. LCCN 2008933269. OCLC 304561132.CS1 maint: ref = harv (связь)
- Андерссон, Мальте (1994). Половой отбор. Монографии по поведению и экологии. Принстон, штат Нью-Джерси: Princeton University Press. ISBN 978-0-691-00057-2. LCCN 93033276. OCLC 28891551.CS1 maint: ref = harv (связь)
- Ариев, Андре (2002). «Платонические и аристотелевские корни телеологических аргументов» (PDF). В Ариев, Андре; Камминс, Роберт; Перлман, Марк (ред.). Функции: Новые очерки философии психологии и биологии. Оксфорд; Нью-Йорк: Oxford University Press. ISBN 978-0-19-824103-4. LCCN 2002020184. OCLC 48965141. Архивировано из оригинал (PDF) 19 февраля 2009 г.CS1 maint: ref = harv (связь)
- Аристотель. Физика. Перевод Р.П. Харди и Р.К. Гей. Архив интернет-классики. OCLC 54350394.CS1 maint: ref = harv (связь)
- Бегон, Майкл; Таунсенд, Колин Р .; Харпер, Джон Л. (1996). Экология: отдельные лица, популяции и сообщества (3-е изд.). Оксфорд; Кембридж, Массачусетс: Blackwell Science. ISBN 978-0-632-03801-5. LCCN 95024627. OCLC 32893848.CS1 maint: ref = harv (связь)
- Кристиансен, Фредди Б. (1984). «Определение и измерение фитнеса». В Shorrocks, Брайан (ред.). Эволюционная экология: 23-й симпозиум Британского экологического общества, Лидс, 1982 г.. Симпозиум Британское экологическое общество. 23. Оксфорд; Бостон: Научные публикации Blackwell. ISBN 978-0-632-01189-6. LCCN 85106855. OCLC 12586581.CS1 maint: ref = harv (связь) Изменено из Кристиансена путем добавления отбора на выживание в репродуктивной фазе.
- Дарвин, Чарльз (1859). О происхождении видов путем естественного отбора или о сохранении избранных рас в борьбе за жизнь (1-е изд.). Лондон: Джон Мюррей. LCCN 06017473. OCLC 741260650.CS1 maint: ref = harv (связь) Книга доступна по адресу Полное собрание сочинений Чарльза Дарвина в Интернете. Проверено 23 июля 2015.
- Дарвин, Чарльз (1861). О происхождении видов путем естественного отбора или о сохранении избранных рас в борьбе за жизнь (3-е изд.). Лондон: Джон Мюррей. LCCN 04001284. OCLC 550913.CS1 maint: ref = harv (связь)
- Дарвин, Чарльз (1872). Происхождение видов посредством естественного отбора или сохранение благоприятных рас в борьбе за жизнь (6-е изд.). Лондон: Джон Мюррей. OCLC 1185571.CS1 maint: ref = harv (связь)
- Дарвин, Чарльз (1958). Барлоу, Нора (ред.). Автобиография Чарльза Дарвина, 1809–1882: восстановлены оригинальные пропуски; Отредактировано и с Приложением и Примечаниями его внучки Норы Барлоу. Лондон: Коллинз. LCCN 93017940. OCLC 869541868.CS1 maint: ref = harv (связь)
- Докинз, Ричард (1976). Эгоистичный ген. Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-857519-1. LCCN 76029168. OCLC 2681149.CS1 maint: ref = harv (связь)
- Деннет, Дэниел С. (1991). Сознание объяснено (1-е изд.). Бостон, Массачусетс: Маленький, коричневый и компания. ISBN 978-0-316-18065-8. LCCN 91015614. OCLC 23648691.CS1 maint: ref = harv (связь)
- Деннет, Дэниел С. (1995). Опасная идея Дарвина: эволюция и смысл жизни. Нью-Йорк: Саймон и Шустер. ISBN 978-0-684-80290-9. LCCN 94049158. OCLC 31867409.CS1 maint: ref = harv (связь)
- Добжанский, Феодосий (1937). Генетика и происхождение видов. Биологические серии Колумбийского университета. Нью-Йорк: Columbia University Press. LCCN 37033383. OCLC 766405.CS1 maint: ref = harv (связь)
- —— (1951). Генетика и происхождение видов. Биологическая серия Колумбийского университета (3-е пересмотренное издание). Нью-Йорк: издательство Колумбийского университета. LCCN 51014816. OCLC 295774.CS1 maint: ref = harv (связь)
- Эйсли, Лорен (1958). Век Дарвина: эволюция и люди, которые ее открыли (1-е изд.). Гарден-Сити, Нью-Йорк: Doubleday. LCCN 58006638. OCLC 168989.CS1 maint: ref = harv (связь)
- Эмпедокл (1898). «Эмпедокл». В Фэрбенкс, Артур (ред.). Первые философы Греции. Перевод Артура Фэрбенкса. Лондон: Kegan Paul, Trench, Trübner & Co. Ltd. LCCN 03031810. OCLC 1376248.CS1 maint: ref = harv (связь) Первые философы Греции на Интернет-архив.
- Эндлер, Джон А. (1986). Естественный отбор в дикой природе. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-08386-5. LCCN 85042683. OCLC 12262762.CS1 maint: ref = harv (связь)
- Энгельс, Фридрих (1964) [1883]. Диалектика природы. 1939 Предисловие J.B.S. Холдейн (3-е изд. Ред.). Москва, СССР: Издатели Progress. LCCN 66044448. OCLC 807047245.CS1 maint: ref = harv (связь) Книга доступна на Марксистский Интернет-архив.
- Фальконер, Дуглас С.; Маккей, Труди F.C. (1996). Введение в количественную генетику (4-е изд.). Харлоу, Англия: Longman. ISBN 978-0-582-24302-6. OCLC 824656731.CS1 maint: ref = harv (связь)
- Фишер, Рональд Эйлмер (1930). Генетическая теория естественного отбора. Оксфорд: Кларендон Пресс. LCCN 30029177. OCLC 493745635.CS1 maint: ref = harv (связь)
- Футуйма, Дуглас Дж. (2005). Эволюция. Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-187-3. LCCN 2004029808. OCLC 57311264.CS1 maint: ref = harv (связь)
- Гольдберг, Дэвид Э. (1989). Генетические алгоритмы в поиске, оптимизации и машинном обучении. Ридинг, Массачусетс: Издательская компания "Эддисон-Уэсли". ISBN 978-0-201-15767-3. LCCN 88006276. OCLC 17674450.CS1 maint: ref = harv (связь)
- Холдейн, Дж. Б.С. (1932). Причины эволюции. Лондон; Нью-Йорк: Longmans, Green & Co. LCCN 32033284. OCLC 5006266.CS1 maint: ref = harv (связь) «Эта книга основана на серии лекций, прочитанных в январе 1931 года в Prifysgol Cymru, Аберистуит, и озаглавлена« Пересмотр дарвинизма ».
- Холдейн, J.B.S. (1954). «Измерение естественного отбора». В Монталенти - Джузеппе; Кьяруги, А. (ред.). Atti del IX Congresso Internazionale di Genetica, Белладжио (Комо) 24–31 назад до 1953 г. [Материалы 9-го Международного конгресса генетиков.]. Кариология. 6 (1953/54) Дополнение. Флоренция, Италия: Университет Флоренции. С. 480–487. OCLC 9069245.CS1 maint: ref = harv (связь)
- Кауфман, Стюарт (1993). Истоки порядка: самоорганизация и отбор в эволюции. Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-507951-7. LCCN 91011148. OCLC 23253930.CS1 maint: ref = harv (связь)
- Ламарк, Жан-Батист (1809). Philosophie Zoologique. Париж: Dentu et L'Auteur. OCLC 2210044.CS1 maint: ref = harv (связь) Philosophie zoologique (1809) на Интернет-архив.
- Лир, Джонатан (1988). Аристотель: желание понять. Кембридж, Великобритания; Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-34762-4. LCCN 87020284. OCLC 16352317.CS1 maint: ref = harv (связь)
- Кун, Томас С. (1996). Структура научных революций (3-е изд.). Чикаго, Иллинойс: Издательство Чикагского университета. ISBN 978-0-226-45808-3. LCCN 96013195. OCLC 34548541.CS1 maint: ref = harv (связь)
- Лемей, Филипп; Салеми, Марко; Vandamme, Anne-Mieke, ред. (2009). Филогенетический справочник: практический подход к филогенетическому анализу и проверке гипотез (2-е изд.). Кембридж, Великобритания; Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-73071-6. LCCN 2009464132. OCLC 295002266.CS1 maint: ref = harv (связь)
- Лукреций (1916). «Книга V». В Леонард, Уильям Эллери (ред.). De rerum natura. Перевод Уильяма Эллери Леонарда. Медфорд / Сомервилл, Массачусетс: Университет Тафтса. OCLC 33233743.CS1 maint: ref = harv (связь)
- Макартур, Роберт Х.; Уилсон, Эдвард О. (2001) [Первоначально опубликовано в 1967 году]. Теория островной биогеографии. Достопримечательности Принстона в биологии. Новое предисловие Эдварда О. Уилсона. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-08836-5. LCCN 00051495. OCLC 45202069.CS1 maint: ref = harv (связь)
- Мальтус, Томас Роберт (1798). Очерк принципа народонаселения, как он влияет на будущее улучшение общества: с замечаниями по поводу предположений г-на Годвина, М. Кондорсе и других авторов (1-е изд.). Лондон: Дж. Джонсон. LCCN 46038215. OCLC 65344349.CS1 maint: ref = harv (связь) Книга есть в наличии здесь от Фрэнка Элвелла, Государственный университет Роджерса.
- Майр, Эрнст (1942). Систематика и происхождение видов с точки зрения зоолога. Колумбийская биологическая серия. 13. Нью-Йорк: издательство Колумбийского университета. LCCN 43001098. OCLC 766053.CS1 maint: ref = harv (связь)
- Майр, Эрнст (2006) [Первоначально опубликовано в 1972 году; Чикаго, Иллинойс: Aldine Publishing Co.]. «Половой отбор и естественный отбор». В Кэмпбелле, Бернард Г. (ред.). Половой отбор и происхождение человека: Дарвиновский поворот. Нью-Брансуик, штат Нью-Джерси: AldineTransaction. ISBN 978-0-202-30845-6. LCCN 2005046652. OCLC 62857839.CS1 maint: ref = harv (связь)
- Мичод, Ричард А. (1999). Дарвиновская динамика: эволюционные переходы в приспособленности и индивидуальности. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-02699-2. LCCN 98004166. OCLC 38948118.CS1 maint: ref = harv (связь)
- Миллер, Джеффри (2000). Брачный разум: как сексуальный выбор повлиял на эволюцию человеческой природы (1-е изд.). Нью-Йорк: Doubleday. ISBN 978-0-385-49516-5. LCCN 00022673. OCLC 43648482.CS1 maint: ref = harv (связь)
- Митчелл, Мелани (1996). Введение в генетические алгоритмы. Сложные адаптивные системы. Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-13316-6. LCCN 95024489. OCLC 42854439.CS1 maint: ref = harv (связь)
- Пинкер, Стивен (1995) [Первоначально опубликовано в 1994; Нью-Йорк: Уильям Морроу и компания ]. Языковой инстинкт: как разум создает язык (1-й Харпер Многолетнее растение ред.). Нью-Йорк: Многолетний Харпер. ISBN 978-0-06-097651-4. LCCN 94039138. OCLC 670524593.CS1 maint: ref = harv (связь)
- Райс, Шон Х. (2004). Эволюционная теория: математические и концептуальные основы. Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-702-8. LCCN 2004008054. OCLC 54988554.CS1 maint: ref = harv (связь)
- Ру, Вильгельм (1881). Der Kampf der Theile im Organismus. Лейпциг: Вильгельм Энгельманн. OCLC 8200805.CS1 maint: ref = harv (связь) Der Kampf der Theile im Organismus на Интернет-архив Проверено 11 августа 2015.
- Трезвый, Эллиотт (1993) [Первоначально опубликовано в 1984 году; Кембридж, Массачусетс: MIT Press]. Природа отбора: эволюционная теория в философском фокусе. Чикаго, Иллинойс: Издательство Чикагского университета. ISBN 978-0-226-76748-2. LCCN 93010367. OCLC 896826726.CS1 maint: ref = harv (связь)
- Уоллес, Альфред Рассел (1871 г.) [Первоначально опубликовано в 1870 г.]. Вклад в теорию естественного отбора. Серия очерков (2-е, с исправлениями и дополнениями ред.). Нью-Йорк: Macmillan & Co. LCCN agr04000394. OCLC 809350209.CS1 maint: ref = harv (связь)
- Уильямс, Джордж К. (1966). Адаптация и естественный отбор: критика некоторых современных эволюционных идей. Научная библиотека Принстона. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-02615-2. LCCN 65017164. OCLC 35230452.CS1 maint: ref = harv (связь)
- Уилсон, Дэвид Слоан (2002). Дарвиновский собор: эволюция, религия и природа общества. Чикаго, Иллинойс: Издательство Чикагского университета. ISBN 978-0-691-02615-2. LCCN 2002017375. OCLC 48777441.CS1 maint: ref = harv (связь)
- Циммер, Карл; Эмлен, Дуглас Дж. (2013). Эволюция: осмысление жизни (1-е изд.). Гринвуд-Виллидж, Колорадо: Робертс и издатели компании. ISBN 978-1-936221-17-2. LCCN 2012025118. OCLC 767565909.CS1 maint: ref = harv (связь)
дальнейшее чтение
- Для технической аудитории
- Белл, Грэм (2008). Выбор: механизм эволюции (2-е изд.). Оксфорд; Нью-Йорк: Oxford University Press. ISBN 978-0-19-856972-5. LCCN 2007039692. OCLC 170034792.
- Джонсон, Клиффорд (1976). Введение в естественный отбор. Балтимор, Мэриленд: University Park Press. ISBN 978-0-8391-0936-5. LCCN 76008175. OCLC 2091640.
- Гулд, Стивен Джей (2002). Структура эволюционной теории. Кембридж, Массачусетс: Belknap Press, издательство Harvard University Press. ISBN 978-0-674-00613-3. LCCN 2001043556. OCLC 47869352.
- Мейнард Смит, Джон (1993) [Первоначально опубликовано в 1958 году; Хармондсворт, Англия: Книги о пингвинах ]. Теория эволюции (Канто ред.). Кембридж, Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-45128-4. LCCN 93020358. OCLC 27676642.
- Поппер, Карл (Декабрь 1978 г.). «Естественный отбор и возникновение разума». Диалектика. 32 (3–4): 339–355. Дои:10.1111 / j.1746-8361.1978.tb01321.x. ISSN 0012-2017.
- Саммут-Бонничи, Таня; Венсли, Робин (сентябрь 2002 г.). «Дарвинизм, вероятность и сложность: рыночные организационные преобразования и изменения, объясненные через теории эволюции» (PDF). Международный журнал обзоров менеджмента. 4 (3): 291–315. Дои:10.1111/1468-2370.00088. ISSN 1460-8545.
- Трезвый, Эллиотт, изд. (1994). Концептуальные вопросы эволюционной биологии (2-е изд.). Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-69162-8. LCCN 93008199. OCLC 28150417.
- Уильямс, Джордж К. (1992). Естественный отбор: области, уровни и проблемы. Оксфордская серия по экологии и эволюции. Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-506933-4. LCCN 91038938. OCLC 228136567.
- Для широкой публики
- Докинз, Ричард (1996). Восхождение на маунт Невероятно (1-е американское изд.). Нью-Йорк: W.W. Нортон и компания. ISBN 978-0-393-03930-6. LCCN 34633422. OCLC 34633422.
- Гулд, Стивен Джей (1977). Со времен Дарвина: размышления в естествознании (1-е изд.). Нью-Йорк: W.W. Нортон и компания. ISBN 978-0-393-06425-4. LCCN 77022504. OCLC 3090189.
- Джонс, Стив (2000). Призрак Дарвина: обновленное происхождение видов (1-е изд.). Нью-Йорк: Случайный дом. ISBN 978-0-375-50103-6. LCCN 99053246. OCLC 42690131.
- Левонтин, Ричард С. (Сентябрь 1978 г.). "Приспособление". Scientific American. 239 (3): 212–230. Bibcode:1978SciAm.239c.212L. Дои:10.1038 / scientificamerican0978-212. ISSN 0036-8733. PMID 705323.
- Майр, Эрнст (2002) [Первоначально опубликовано в 2001; Нью-Йорк: Базовые книги ]. Что такое эволюция. Мастера наук. Лондон: Вайденфельд и Николсон. ISBN 978-0-297-60741-0. LCCN 2001036562. OCLC 248107061.
- Вайнер, Джонатан (1994). Клюв вьюрка: история эволюции в наше время (1-е изд.). Нью-Йорк: Knopf. ISBN 978-0-679-40003-5. LCCN 93036755. OCLC 29029572.
- Исторический
- Кон, Марек (2004). Причина всему: естественный отбор и английское воображение. Лондон: Фабер и Фабер. ISBN 978-0-571-22392-3. LCCN 2005360890. OCLC 57200626.
- Циркле, Конвей (25 апреля 1941 г.). "Естественный отбор до" происхождения видов "'". Труды Американского философского общества. 84 (1): 71–123. ISSN 0003-049X. JSTOR 984852.
внешняя ссылка
- Дарвин, Чарльз. «О происхождении видов». Архивировано из оригинал 25 февраля 2001 г. - Глава 4, Естественный отбор
