Орбитофронтальная кора - Orbitofrontal cortex
| Орбитофронтальная кора | |
|---|---|
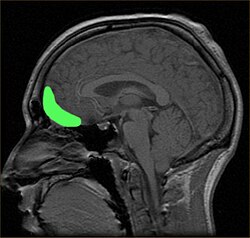 Примерное расположение ОФК показано на сагиттальной МРТ | |
 Глазничная поверхность левой лобной доли. | |
| Подробности | |
| Часть | Лобная доля |
| Идентификаторы | |
| латинский | орбитофронтальная кора |
| NeuroNames | 91 |
| НейроЛекс Я БЫ | birnlex_1049 |
| FMA | 242003 |
| Анатомические термины нейроанатомии | |
В орбитофронтальная кора (OFC) это префронтальная кора регион в фронтальные доли из мозг который участвует в когнитивный процесс из принимать решение. У нечеловеческих приматов он состоит из ассоциативные области коры Площадь Бродмана 11, 12 и 13; у людей он состоит из Площадь Бродмана 10, 11 и 47.[1]
OFC считается анатомически синонимом вентромедиальная префронтальная кора.[2] Следовательно, этот регион отличается отчетливыми нейронными связями и различными функциями, которые он выполняет.[3] Он определяется как часть префронтальной коры, которая получает проекции от медиальное дорсальное ядро из таламус, и считается, что он представляет эмоция и награда в принимать решение.[4] Он получил свое название от своего положения непосредственно над орбиты в которой глаза расположены. В ОФК человека обнаружена значительная индивидуальная изменчивость.[5] Связанная область находится в грызуны.[6]
Структура
OFC разделен на несколько широких областей, различающихся цитоархитектурой, включая площадь Бродмана 47/12, площадь Бродмана 11, площадь Бродмана 14, площадь Бродмана 13, и площадь Бродмана 10.[7] Четыре извилины разделены комплексом борозд, который чаще всего напоминает узор "H" или "K". Две борозды, латеральная и глазничная, проходят вдоль ростро-каудальной оси и обычно соединяются поперечной орбитальной бороздой, которая проходит вдоль медиально-боковой оси. Медиально медиальная орбитальная извилина отделена от прямая извилина обонятельной бороздой.[8] Спереди как прямая извилина, так и медиальная часть медиальной орбитальной извилины состоят из области 11 (м), а сзади - области 14. Задняя глазничная извилина состоит в основном из области 13 и ограничена медиально и латерально передними конечностями медиальная и латеральная борозды глазницы. Область 11 составляет большую часть OFC, включая латеральные части медиальной орбитальной извилины, а также переднюю глазничную извилину. Боковая орбитальная извилина состоит преимущественно из области 47/12.[7] Большая часть OFC гранулированный, хотя хвостовые части области 13 и области 14 агранулярные.[9] Эти каудальные области, которые иногда включают части островковая кора, реагирует в первую очередь на необработанные сенсорные сигналы.[10]
Подключения
Связность OFC несколько варьируется вдоль рострально-каудальной оси. Хвостовая часть OFC в большей степени связана с сенсорными областями, в частности, получает прямой сигнал от грушевидная кора. Хвостовая часть OFC также наиболее тесно связана с миндалина.[11] Рострально OFC получает меньше прямых сенсорных проекций и меньше связан с миндалевидным телом, но он взаимосвязан с боковой префронтальной корой и парагиппокамп.[10] Связность OFC также была концептуализирована как состоящая из двух сетей; орбитальная сеть, состоящая из большинства центральных частей OFC, включая большую часть областей 47/12, 13 и 11; медиальная сеть, состоящая из самой медиальной и каудолатеральной областей OFC, а также областей 24, 25 и 32 медиальной префронтальной коры.[12] Медиальную и орбитальную сети иногда называют «висцеромоторной сетью» и «сенсорной сетью» соответственно.[13]
Афференты
OFC получает проекции от множества сенсорных модальностей. В первичная обонятельная кора, вкусовая кора, вторичная соматосенсорная кора, начальство и нижняя височная извилина (передача визуальной информации) весь проект в OFC.[9][14][15] Доказательства слуховых входов слабы, хотя некоторые нейроны реагируют на слуховые стимулы, указывая на то, что существует косвенная проекция.[12] OFC также получает информацию от медиальное дорсальное ядро, островковая кора, энторинальная кора, периринальная кора, гипоталамус, и миндалина.[14][16]
Efferents
Орбитофронтальная кора реципрокно связана с периринальной и энторинальной корой,[16] миндалевидное тело, гипоталамус и части медиальной височной доли. В дополнение к этим результатам, OFC также планирует полосатое тело, в том числе прилежащее ядро, хвостатое ядро, и вентральный скорлупа, а также области среднего мозга, включая периакведуктальный серый, и вентральная тегментальная область.[14][17] Входы OFC в синапс миндалевидного тела по нескольким мишеням, включая два надежных пути к базолатеральная миндалина и интеркалированные клетки миндалины, а также более слабая прямая проекция на центральное ядро миндалины.[11]
Функция
OFC было возложено несколько функций, включая посредничество при реагировании на контекст,[18] гибкое кодирование непредвиденных обстоятельств, кодирование значения, кодирование предполагаемого значения, подавление реакции, обучение изменениям в непредвиденных обстоятельствах, эмоциональная оценка,[19] изменение поведения с помощью соматических маркеров, управление социальным поведением и представление пространств состояний.[20][21] Хотя большинство этих теорий объясняют определенные аспекты электрофизиологических наблюдений и изменения поведения, связанные с поражением, они часто не могут объяснить или опровергаются другими выводами. Одно из предложений, объясняющее разнообразие функций OFC, состоит в том, что OFC кодирует пространства состояний или дискретную конфигурацию внутренних и внешних характеристик, связанных с ситуацией и ее непредвиденными обстоятельствами.[22] Например, предложение о том, что OFC кодирует экономическую ценность, может быть отражением значения состояния задачи кодирования OFC.[18] Представление состояний задач может также объяснить предложение о том, что OFC действует как гибкая карта непредвиденных обстоятельств, поскольку переключение состояния задачи позволит кодировать новые непредвиденные обстоятельства в одном состоянии с сохранением старых непредвиденных обстоятельств в отдельном состоянии, что позволяет переключение непредвиденных обстоятельств, когда старое состояние задачи снова становится актуальным.[21] Представление состояний задачи подтверждается электрофизиологическими данными, демонстрирующими, что OFC реагирует на разнообразный набор функций задачи и может быстро переназначаться во время смены непредвиденных обстоятельств.[21] Представление состояний задач может влиять на поведение через множество потенциальных механизмов. Например, OFC необходим для вентральная тегментальная область (ВТА) нейроны, чтобы произвести ошибку предсказания дофаминергического вознаграждения, и OFC может кодировать ожидания для вычисления RPE в VTA.[18]
Конкретные функции были возложены на субрегионы OFC. Боковой OFC был предложен для отражения потенциальной ценности выбора, позволяя фиктивным (контрфактическим) ошибкам прогнозирования потенциально опосредовать выбор переключения во время разворота, исчезновения и девальвации.[23] Оптогенетическая активация lOFC усиливает цель, направленную над привычным поведением, возможно, отражая повышенную чувствительность к потенциальному выбору и, следовательно, повышенное переключение. С другой стороны, mOFC было предложено для отражения относительной субъективной ценности.[19] У грызунов аналогичная функция была приписана mOFC, кодирующей значение действия градуированным образом, в то время как lOFC был предложен для кодирования определенных сенсорных характеристик результатов.[24] LOFC также был предложен для кодирования ассоциаций результатов стимула, которые затем сравниваются по значению в mOFC.[25] Мета-анализ нейровизуализационных исследований на людях показывает, что существует медиально-латеральный градиент валентности, при этом медиальный OFC чаще всего реагирует на вознаграждение, а латеральный OFC чаще всего отвечает на наказание. Также был обнаружен задне-передний градиент абстрактности, при этом задний OFC реагировал на более простое вознаграждение, а передний OFC более отвечал на абстрактное вознаграждение.[26] Аналогичные результаты были получены в метаанализе исследований первичного и вторичного вознаграждения.[27]
OFC и базолтеральная миндалина (BLA) сильно взаимосвязаны, и их взаимосвязь необходима для задач девальвации. Повреждение либо BLA, либо OFC ранее, но только OFC после девальвации ухудшает производительность.[28] В то время как BLA реагирует только на реплики, предсказывающие значимые исходы дифференцированным образом в соответствии со значением, OFC реагирует как на ценность, так и на специфические сенсорные атрибуты ассоциаций реплика-результат. В то время как нейроны OFC, которые на ранних этапах обучения реагируют на получение результата, обычно передают свою реакцию на появление сигналов, которые предсказывают результат, повреждение BLA ухудшает эту форму обучения.[29]
Задняя орбитофронтальная кора головного мозга (pOFC) связана с миндалевидным телом несколькими путями, которые способны как повышать, так и понижать активность вегетативной нервной системы.[30] Предварительные данные свидетельствуют о том, что нейромодулятор дофамин играет роль в обеспечении баланса между тормозными и возбуждающими путями, с высоким уровнем дофамина. автономный Мероприятия.[31]
Было высказано предположение, что медиальный OFC участвует в создании ассоциаций стимул-вознаграждение и в подкреплении поведения, в то время как латеральный OFC участвует в ассоциациях стимул-результат, а также в оценке и, возможно, изменении поведения.[32] Активность в боковом OFC обнаруживается, например, когда субъекты кодируют новые ожидания относительно наказания и социальных репрессалий.[33][34]
Было обнаружено, что средне-передний OFC постоянно отслеживает субъективные удовольствие в исследованиях нейровизуализации. А гедонистическая точка доступа был обнаружен в передней части OFC, которая способна усиливать ответную реакцию на сахарозу. OFC также способен искажать аффективные реакции, вызванные α-амино-3-гидрокси-5-метил-4-изоксазолепропионовая кислота (AMPA) антагонизм в прилежащее ядро к аппетитным ответам.[35]
OFC способен модулировать агрессивное поведение через проекции на интернейроны в миндалине, которые подавляют глутаминергические проекции на вентромедиальные клетки. гипоталамус.[36]
Электрофизиология
Нейроны в OFC реагируют как на первичные подкрепления, так и на сигналы, предсказывающие вознаграждение в нескольких сенсорных областях. Доказательства реакций на зрительные, вкусовые, соматосенсорные и обонятельные раздражители надежны, но доказательства для слуховых реакций слабее. В подмножестве нейронов OFC нейронные реакции на награды или сигналы вознаграждения модулируются индивидуальными предпочтениями и внутренними мотивационными состояниями, такими как голод. Часть нейронов, которые реагируют на сенсорные сигналы, предсказывающие вознаграждение, избирательны в отношении вознаграждения и демонстрируют обратное поведение, когда отношения исходных сигналов меняются местами. Нейроны в OFC также реагируют на отсутствие ожидаемого вознаграждения и наказания. Другая популяция нейронов реагирует на новые стимулы и может «запоминать» знакомые стимулы до суток.[37]
Во время выполнения заданий на вознаграждение или инструментальное вознаграждение нейроны в OFC демонстрируют три основных типа срабатывания; стрельба в ответ на реплики; стрельба до получения награды; стрельба в ответ на получение награды. В отличие от медиальной префронтальной коры и полосатое тело, Нейроны OFC не проявляют возбуждения, опосредованного движением. Однако их предсказательные ответы на вознаграждение формируются вниманием: при переключении внимания между двумя альтернативами одна и та же популяция OFC будет положительно представлять ценность текущего предмета, но отрицательно - ценность оставленного без присмотра предмета.[38] Кодирование величины вознаграждения также является гибким и учитывает относительные значения существующих вознаграждений.[39]
Люди
OFC человека - одна из наименее изученных областей человеческого мозга; но было высказано предположение, что OFC участвует в сенсорной интеграции, в представлении аффективной ценности подкреплений и в принимать решение и ожидание.[1] В частности, OFC, по-видимому, играет важную роль в сигнализировании об ожидаемых наградах / наказаниях за действие с учетом конкретных деталей ситуации.[40] При этом мозг способен сравнивать ожидаемую награду / наказание с фактической доставкой награды / наказания, что делает OFC критически важным для адаптивного обучения. Это подтверждается исследованиями на людях, приматах и грызунах.
Психиатрические расстройства
Орбитофронтальная кора головного мозга участвует в пограничное расстройство личности,[41] шизофрения, сильное депрессивное расстройство, биполярное расстройство, обсессивно-компульсивное расстройство, зависимость, пост-травматическое стрессовое растройство, Аутизм,[42] и паническое расстройство. Хотя нейровизуализационные исследования предоставили доказательства дисфункции при широком спектре психических расстройств, загадочная природа роли ОФК в поведении усложняет понимание их роли в патофизиологии психических расстройств.[43] Функция OFC неизвестна, но его анатомические связи с вентральным полосатым телом, миндалевидным телом, гипоталамусом, гиппокампом и периакведуктальным серым цветом поддерживают роль посредника в поведении, связанном с вознаграждением и страхом.[44]
Обсессивно-компульсивное расстройство
Мета-анализы нейровизуализационных исследований при ОКР показывают гиперактивность в областях, которые обычно считаются частью орбитофронтального сегмента кортико-базальные ганглии-таламо-кортикальная петля такой как хвостатое ядро, таламус и орбитофронтальная кора. Было высказано предположение, что ОКР отражает положительную обратную связь из-за взаимного возбуждения OFC и подкорковых структур.[45] В то время как OFC обычно сверхактивен во время задач по провокации симптомов, когнитивные задачи обычно вызывают гипоактивность OFC;[46] это может отражать различие между эмоциональными и неэмоциональными задачами, латеральными и медиальными OFC,[47] или просто несовместимые методологии.[48]
Зависимость
Модели на животных и клеточно-специфические манипуляции в отношении поведения, связанного с поиском наркотиков, указывают на нарушение функции OFC в зависимости.[49] Расстройства, связанные с употреблением психоактивных веществ связаны с различными недостатками, связанными с гибким целенаправленным поведением и принятием решений. Эти дефициты частично совпадают с симптомами, связанными с поражениями OFC, а также связаны с уменьшением орбитофронтальной серое вещество, гипометаболизм в состоянии покоя и притупление активности ОФК во время выполнения задач, связанных с принятием решений или целенаправленным поведением. В отличие от состояния покоя и активности, связанной с принятием решений, сигналы, связанные с наркотиками, вызывают устойчивую активность OFC, которая коррелирует с влечением.[50] Исследования грызунов также демонстрируют, что проекции lOFC на BLA необходимы для восстановления самостоятельного введения, вызванного сигналом. Все эти результаты согласуются с ролью, которую OFC играет в кодировании результатов, связанных с определенными стимулами.[51][52][53] Прогресс в сторону компульсивного злоупотребления психоактивными веществами может отражать переход от принятия решений на основе модели, где решения определяют внутренняя модель будущих результатов, к моделированию свободного обучения, где решения основываются на истории подкрепления. Обучение на основе моделей включает в себя OFC и является гибким и целенаправленным, в то время как обучение на основе моделей более жесткое; поскольку переход к более модельному свободному поведению из-за дисфункции в OFC, например, вызванного злоупотреблением наркотиками, может лежать в основе привычек поиска наркотиков.[54]
Поведенческие расстройства
Расстройство поведения связано как со структурными аномалиями, так и с функциональными аномалиями при выполнении аффективных задач.[55] Нарушения в структуре, активности и функциональной связности ОФК наблюдались в связи с агрессией.[56]
Аффективные расстройства
Нейровизуализационные исследования выявили аномалии OFC как при БДР, так и при биполярном расстройстве. В соответствии с градиентом медиального / вознаграждения и бокового / наказательного, обнаруженного в исследованиях нейровизуализации, некоторые исследования нейровизуализации наблюдали повышенную латеральную активность OFC при депрессии, а также снижение взаимосвязанности медиальных OFC и усиление взаимосвязи в латеральных OFC.[57] Гипоактивность боковых ОФК часто наблюдается при биполярном расстройстве, особенно во время маниакальных эпизодов.[57]
Исследование
Изображения
С помощью функциональная магнитно-резонансная томография (фМРТ) для изображения OFC человека является сложной задачей, потому что эта область мозга находится в непосредственной близости от наполненные воздухом пазухи. Это означает, что ошибки артефактов может произойти в обработка сигналов, вызывая, например, геометрические искажения, которые обычны при использовании эхопланарное изображение (EPI) при более высокой напряженности магнитного поля. Поэтому рекомендуется проявлять особую осторожность для получения хорошего сигнала от орбитофронтальной коры, и был разработан ряд стратегий, таких как автоматическое шимминг при высоких значениях статического магнитного поля.[58]
Грызунов
В грызуны, OFC полностью агранулярный или дисгранулярный.[9] OFC делится на вентролатеральную (VLO), латеральную (LO), медиальную (MO) и дорсолатеральную (DLO) области.[12] Использование высокоспециализированных методов для управления схемами, таких как оптогенетика, OFC был вовлечен в поведение, подобное ОКР[59], а также в умении использовать скрытые переменные в задаче принятия решения[60].
Клиническое значение
Повреждать
Разрушение ОФК через приобретенная травма головного мозга обычно приводит к модели расторможенный поведение. Примеры включают чрезмерную ругань, гиперсексуальность, плохое социальное взаимодействие, компульсивные азартные игры, употребление наркотиков (включая алкоголь и табак) и плохую способность к сочувствию. Расторможенное поведение пациентов с некоторыми формами лобно-височная деменция считается, что это вызвано дегенерацией OFC.[61]
Нарушение
Когда соединение OFC нарушается, может возникнуть ряд когнитивных, поведенческих и эмоциональных последствий. Исследования подтверждают, что основные расстройства, связанные с нерегулируемыми связями / схемами OFC, связаны с принятием решений, регулированием эмоций и ожиданием вознаграждения.[62][63][64] Недавнее мультимодальное исследование нейровизуализации человека показывает нарушение структурной и функциональной связи OFC с подкорковыми лимбическими структурами (например, миндалевидным телом или гиппокампом) и другими лобными областями (например, дорсальной префронтальной корой или передней поясной корой) коррелирует с аномальным влиянием OFC ( например, страх) обработка у клинически тревожных взрослых.[65]
Одним из явных продолжений проблем с принятием решений является наркомания /зависимость от веществ, что может быть результатом нарушения стриато-таламо-орбитофронтальной цепи.[64][62][66] Синдром дефицита внимания и гиперактивности (СДВГ) также был замешан в дисфункции нейронной схемы вознаграждения, контролирующей мотивацию, вознаграждение и импульсивность, включая системы OFC.[63] Другие нарушения исполнительной функции и импульсный контроль могут быть затронуты нарушением регуляции схемы OFC, например обсессивно-компульсивное расстройство и трихотилломания[67][68][69]
Некоторые виды деменции также связаны с нарушениями связи OFC. Поведенческий вариант лобно-височная деменция[70] ассоциируется с паттернами нейральной атрофии проекционных волокон белого и серого вещества, участвующих в связности OFC.[71] Наконец, некоторые исследования показывают, что на более поздних стадиях Болезнь Альцгеймера будут затронуты измененной связью систем OFC.[69]
Орбитофронтальная эпилепсия
Орбитофронтальная эпилепсия встречается редко, но встречается. Проявления OFC эпилепсии довольно разнообразны, хотя общие характеристики включают связанные со сном, автоматизм и гипермоторные симптомы. В одном обзоре сообщалось, что ауры, как правило, не были обычными или неспецифическими, в то время как в другом сообщалось, что с эпилепсией OFC связаны ауры, связанные с соматосенсорным феноменом и страхом.[72][73][74]
Оценка
Тест визуальной дискриминации
Тест визуальной дискриминации состоит из двух компонентов. В первом компоненте, «обратное обучение», участникам предоставляется одно из двух изображений, A и B. Они узнают, что они будут вознаграждены, если нажмут кнопку, когда отображается изображение A, но будут наказаны, если они нажмут кнопку, когда изображение Отображается B. Как только это правило установлено, оно меняется. Другими словами, теперь правильно нажимать кнопку для изображения B, а не для изображения A. Большинство здоровых участников почти сразу усваивают изменение этого правила, но пациенты с повреждением OFC продолжают реагировать на исходный шаблон подкрепления, хотя они теперь его наказывают за то, что он продолжал с этим. Rolls et al.[75] отметили, что этот образец поведения особенно необычен, учитывая, что пациенты сообщили, что они поняли правило.
Вторая составляющая теста - «угасание». Опять же, участники учатся нажимать кнопку для изображения A, но не для изображения B. Однако на этот раз вместо изменения правил правила полностью меняются. Теперь участник будет наказан за нажатие кнопки в ответ на либо рисунок. Правильный ответ - вообще не нажимать кнопку, но людям с дисфункцией OFC трудно устоять перед соблазном нажать кнопку, несмотря на то, что они за это наказаны.
Iowa Gambling Task
Симуляция реальной жизни принимать решение, то Задание на азартные игры в Айове широко используется в исследованиях познания и эмоций.[76] Участникам представлены четыре виртуальные колоды карт на экране компьютера. Им говорят, что каждый раз, когда они выбирают карту, они выигрывают игровые деньги. Им говорят, что цель игры - выиграть как можно больше денег. Однако время от времени, выбирая карту, они теряют немного денег. Задача должна быть непрозрачной, то есть участники не предназначены для сознательной выработки правила, и они должны выбирать карты на основе их "инстинктивная реакция. »Две из колод являются« плохими колодами », что означает, что в течение достаточно длительного времени они будут приносить чистый убыток; другие две колоды являются« хорошими колодами »и со временем принесут чистый выигрыш.
Большинство здоровых участников пробуют карты из каждой колоды, и после примерно 40 или 50 выборов довольно хорошо умеют придерживаться хороших колод. Однако пациенты с дисфункцией OFC продолжают настойчиво с плохими колодами, иногда даже если они знают, что в целом теряют деньги. Параллельное измерение кожно-гальваническая реакция показывает, что здоровые участники демонстрируют «стрессовую» реакцию на зависание над бэд-деками всего после 10 попыток, задолго до осознанного ощущения, что деки плохие. Напротив, у пациентов с дисфункцией ОФК никогда не развивается эта физиологическая реакция на надвигающееся наказание. Бечара и его коллеги объясняют это с точки зрения гипотеза соматического маркера. Задача по азартным играм в Айове в настоящее время используется рядом исследовательских групп, использующих фМРТ изучить, какие области мозга активируются задачей у здоровых добровольцев, а также в клинических группах с такими состояниями, как шизофрения и обсессивно-компульсивное расстройство.
Тест на оплошность - это серия виньеток, рассказывающих о социальном событии, во время которого кто-то сказал что-то, о чем не следовало говорить, или о неприятном происшествии. Задача участника - определить, что было сказано, что было неловко, почему это было неловко, как люди бы отреагировали на оплошность и на контрольный вопрос. Хотя изначально он был разработан для использования людьми на спектр аутизма,[77] Тест также чувствителен к пациентам с дисфункцией OFC, которые не могут судить о том, когда произошло что-то социально неудобное, несмотря на то, что они, кажется, прекрасно понимают историю.
Смотрите также
Дополнительные изображения
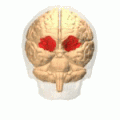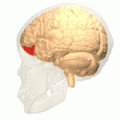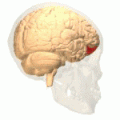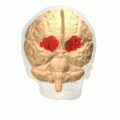
Орбитальная извилина показана красным.

Медиальная поверхность коры головного мозга - извилины

Базальная поверхность головного мозга. Орбитальная извилина показана красным цветом.

Боковая орбитофронтальная кора

Медиальная орбитофронтальная кора, вид внутреннего среза.





Рекомендации
- ^ а б Крингельбах М. Л. (2005). «Орбитофронтальная кора: связь награды с гедонистическим опытом». Обзоры природы Неврология. 6 (9): 691–702. Дои:10.1038 / nrn1747. PMID 16136173.
- ^ Филлипс, LH., Макферсон, SE. И Делла Сала, С. (2002). «Возраст, познание и эмоции: роль анатомической сегрегации в лобных долях: роль анатомической сегрегации в лобных долях». в J Grafman (ed.), Справочник по нейропсихологии: лобные доли. Elsevier Science, Амстердам, стр. 73-98.
- ^ Barbas H, Ghashghaei H, Rempel-Clower N, Xiao D (2002) Анатомические основы функциональной специализации префронтальной коры приматов. В: Справочник по нейропсихологии (Grafman J, ed), стр. 1-27. Амстердам: Elsevier Science B.V.
- ^ Фустер, Дж. Префронтальная кора, (Raven Press, Нью-Йорк, 1997).
- ^ Исама Н., Фейсон В., Пейн М.Э., Макфолл Дж., Стеффенс Д.К., Бейер Дж. Л., Кришнан Р., Тейлор В. Д. (2010). «Вариабельность лобно-височной структуры мозга: важность привлечения афроамериканцев в нейробиологические исследования». PLOS ONE. 5 (10): e13642. Bibcode:2010PLoSO ... 513642I. Дои:10.1371 / journal.pone.0013642. ЧВК 2964318. PMID 21049028.
- ^ Uylings HB, Groenewegen HJ, Kolb B (2003). «Есть ли у крыс префронтальная кора?». Behav Brain Res. 146 (1–2): 3–17. Дои:10.1016 / j.bbr.2003.09.028. PMID 14643455.
- ^ а б Макки, Сотт; Петридес, Майкл (2006). «Глава 2: Орбитофронтальная кора: морфология и архитектура борозды и гирали». В Zald, David H .; Раух, Скотт (ред.). Орбитофронтальная кора. Нью-Йорк: Издательство Оксфордского университета. п. 34. ISBN 9780198565741.
- ^ Макки, Сотт; Петридес, Майкл (2006). «Глава 2: Орбитофронтальная кора: морфология и архитектура борозды и гирали». В Zald, David H .; Раух, Скотт (ред.). Орбитофронтальная кора. Нью-Йорк: Издательство Оксфордского университета. п. 24. ISBN 9780198565741.
- ^ а б c Passingham, Richard E .; Мудрый, Стивен П. (1012). «Глава 4 Орбитальная префронтальная кора: выбор объектов на основе результатов». Нейробиология префронтальной коры: анатомия, эволюция и происхождение понимания. Грейт-Кларендон-стрит, Оксфорд: Издательство Оксфордского университета. п. 97. ISBN 9780199552917.
- ^ а б Haber, SN; Behrens, TE (3 сентября 2014 г.). «Нейронная сеть, лежащая в основе обучения, основанного на стимулах: значение для интерпретации сбоев в цепи при психических расстройствах». Нейрон. 83 (5): 1019–39. Дои:10.1016 / j.neuron.2014.08.031. ЧВК 4255982. PMID 25189208.
- ^ а б Барбас, Елена; Зикопулос, Василис (2006). «Глава 4: Последовательные и параллельные схемы для обработки эмоций в орбитофронтальной коре приматов». В Rauch, Scott L .; Зальд, Дэвид Х. (ред.). Орбитофронтальная кора. Нью-Йорк: Издательство Оксфордского университета. п. 67.
- ^ а б c Прайс, Джозеф Л. (2006). «Глава 3: Связи орбитальной коры». В Rauch, Scott L .; Зальд, Дэвид Х. (ред.). Орбитофронтальная кора. Нью-Йорк: Издательство Оксфордского университета. п. 42.
- ^ Рудебек, PH; Мюррей, EA (декабрь 2011 г.). «Балканизация орбитофронтальной коры приматов: отдельные субрегионы для сравнения и сопоставления значений». Летопись Нью-Йоркской академии наук. 1239 (1): 1–13. Bibcode:2011НЯСА1239 .... 1Р. Дои:10.1111 / j.1749-6632.2011.06267.x. ЧВК 3951748. PMID 22145870.
- ^ а б c Rolls, ET (март 2000 г.). «Орбитофронтальная кора и награда». Кора головного мозга. 10 (3): 284–94. Дои:10.1093 / cercor / 10.3.284. PMID 10731223.
- ^ Rolls, ET (ноябрь 2004 г.). «Конвергенция сенсорных систем в орбитофронтальной коре у приматов и дизайн мозга для эмоций». Анатомическая запись, часть A: открытия в молекулярной, клеточной и эволюционной биологии. 281 (1): 1212–25. Дои:10.1002 / ar.a.20126. PMID 15470678.
- ^ а б Ремпель-Клауэр, Нидерланды (декабрь 2007 г.). «Роль орбитофронтальных связей коры в эмоциях». Летопись Нью-Йоркской академии наук. 1121 (1): 72–86. Bibcode:2007НЯСА1121 ... 72Р. Дои:10.1196 / летопись.1401.026. PMID 17846152.
- ^ Прайс, Джозеф Л. (2006). «Глава 3: Связи орбитальной коры». В Rauch, Scott L .; Зальд, Дэвид Х. (ред.). Орбитофронтальная кора. Нью-Йорк: Издательство Оксфордского университета. п. 45.
- ^ а б c Викенхайзер, AM; Шенбаум, G (август 2016 г.). «За рекой, через лес: когнитивные карты в гиппокампе и орбитофронтальной коре». Обзоры природы. Неврология. 17 (8): 513–23. Дои:10.1038 / номер 2016.56. ЧВК 5541258. PMID 27256552.
- ^ а б Fettes, P; Шульце, Л; Даунар, Дж (2017). "Кортико-стриатально-таламические петли орбитофронтальной коры: многообещающие терапевтические цели при психических заболеваниях". Границы системной нейробиологии. 11: 25. Дои:10.3389 / fnsys.2017.00025. ЧВК 5406748. PMID 28496402.
- ^ Уилсон, Роберт С .; Takahashi, Yuji K .; Шенбаум, Джеффри; Нив, Яэль (январь 2014). «Орбитофронтальная кора как когнитивная карта пространства задач». Нейрон. 81 (2): 267–279. Дои:10.1016 / j.neuron.2013.11.005. ISSN 0896-6273. ЧВК 4001869. PMID 24462094.
- ^ а б c Sadacca, BF; Викенхайзер, AM; Шенбаум, Г. (14 марта 2017 г.). «К теоретической роли тонического норадреналина в орбитофронтальной коре головного мозга в облегчении гибкого обучения». Неврология. 345: 124–129. Дои:10.1016 / j.neuroscience.2016.04.017. ЧВК 5461826. PMID 27102419.
- ^ Стальнакер Т.А.; Cooch, NK; Шенбаум, Г. (май 2015 г.). «Чего не делает орбитофронтальная кора». Природа Неврология. 18 (5): 620–7. Дои:10.1038 / № 3982. ЧВК 5541252. PMID 25919962.
- ^ Tobia, M. J .; Guo, R .; Schwarze, U .; Boehmer, W .; Gläscher, J .; Finckh, B .; Marschner, A .; Büchel, C .; Obermayer, K .; Соммер, Т. (2014-04-01). «Нейронные системы для выбора и оценки с контрфактическими обучающими сигналами». NeuroImage. 89: 57–69. Дои:10.1016 / j.neuroimage.2013.11.051. ISSN 1053-8119. PMID 24321554.
- ^ Искьердо, А (1 ноября 2017 г.). «Функциональная неоднородность орбитофронтальной коры головного мозга крысы в поощрении обучения и принятия решений». Журнал неврологии. 37 (44): 10529–10540. Дои:10.1523 / JNEUROSCI.1678-17.2017. ЧВК 6596524. PMID 29093055.
- ^ Рудебек, PH; Мюррей, EA (декабрь 2011 г.). «Балканизация орбитофронтальной коры приматов: отдельные субрегионы для сравнения и сопоставления значений». Летопись Нью-Йоркской академии наук. 1239 (1): 1–13. Bibcode:2011НЯСА1239 .... 1Р. Дои:10.1111 / j.1749-6632.2011.06267.x. ЧВК 3951748. PMID 22145870.
- ^ Крингельбах, ML; Rolls, ET (апрель 2004 г.). «Функциональная нейроанатомия орбитофронтальной коры человека: данные нейровизуализации и нейропсихологии». Прогресс в нейробиологии. 72 (5): 341–72. Дои:10.1016 / j.pneurobio.2004.03.006. PMID 15157726.
- ^ Сескус, G; Caldú, X; Сегура, Б; Дреер, JC (май 2013 г.). «Обработка первичных и вторичных вознаграждений: количественный мета-анализ и обзор исследований функциональной нейровизуализации человека». Неврология и биоповеденческие обзоры. 37 (4): 681–96. Дои:10.1016 / j.neubiorev.2013.02.002. HDL:2066/117487. PMID 23415703.
- ^ Padoa-Schioppa, C; Конен, KE (15 ноября 2017 г.). «Орбитофронтальная кора: нейронный контур для принятия экономических решений». Нейрон. 96 (4): 736–754. Дои:10.1016 / j.neuron.2017.09.031. ЧВК 5726577. PMID 29144973.
- ^ Шарп, MJ; Шенбаум, G (май 2016 г.). «Назад к основам: прогнозирование орбитофронтально-миндалевидного контура». Нейробиология обучения и памяти. 131: 201–6. Дои:10.1016 / j.nlm.2016.04.009. ЧВК 5541254. PMID 27112314.
- ^ Барбас, H (август 2007 г.). «Поток информации об эмоциях через височные и орбитофронтальные пути». Журнал анатомии. 211 (2): 237–49. Дои:10.1111 / j.1469-7580.2007.00777.x. ЧВК 2375774. PMID 17635630.
Задняя орбитофронтальная кора нацелена на двойные системы миндалевидного тела, которые оказывают противоположное влияние на центральные вегетативные структуры. Оба пути берут начало в задней орбитофронтальной коре, но один из них сильно нацелен на тормозящие интеркалированные массы, активация которых может в конечном итоге подавить центральные вегетативные структуры во время эмоционального возбуждения.
- ^ Зикопулос, В; Höistad, M; Джон, Y; Barbas, H (17 мая 2017 г.). «Задние орбитофронтальные и передние поясные пути к тормозящей и возбуждающей системам миндалины с противоположными функциями». Журнал неврологии. 37 (20): 5051–5064. Дои:10.1523 / JNEUROSCI.3940-16.2017. ЧВК 5444191. PMID 28411274.
Обнаруженная здесь специфическая иннервация тормозных систем в миндалевидном теле, наряду с дифференциальным воздействием на них дофамина, позволяет предположить, как могут быть достигнуты различные вегетативные состояния. Сильное влияние pOFC на IM, которое активирует нейроны DARPP-32 + и CB +, может помочь модулировать вегетативную функцию за счет подавления CeM и тем самым облегчить социальные взаимодействия у приматов ... С другой стороны, в состоянии паники, когда выживание воспринимается как под угрозой, уровень дофамина заметно повышается. Таким образом, нейроны DARPP-32 + в IM могут быть главным образом ингибированы, что делает путь pOFC неэффективным.
- ^ Уолтон М. Э .; Беренс Т. Э .; Бакли М. Дж .; Rudebeck P. H .; Рашворт М. Ф. (2010). «Разделимые системы обучения в головном мозге макака и роль орбитофронтальной коры в случайном обучении». Нейрон. 65 (6): 927–939. Дои:10.1016 / j.neuron.2010.02.027. ЧВК 3566584. PMID 20346766.
- ^ Campbell-Meiklejohn D. K .; Kanai R .; Bahrami B .; Бах Д. Р .; Долан Р. Дж .; Roepstorff A .; Фрит К. Д. (2012). «Структура орбитофронтальной коры предсказывает социальное влияние». Текущая биология. 22 (4): R123 – R124. Дои:10.1016 / j.cub.2012.01.012. ЧВК 3315000. PMID 22361146.
- ^ Tanferna A .; Лопес-Хименес Л .; Blas J .; Хиральдо Ф .; Серджио Ф. (2012). «Как совет эксперта влияет на принятие решений». PLoS ONE. 7 (11): e49748. Bibcode:2012PLoSO ... 749748M. Дои:10.1371 / journal.pone.0049748. ЧВК 3504100. PMID 23185425.
- ^ Берридж, KC; Крингельбах, ML (6 мая 2015 г.). «Системы удовольствия в мозгу». Нейрон. 86 (3): 646–64. Дои:10.1016 / j.neuron.2015.02.018. ЧВК 4425246. PMID 25950633.
- ^ Нуман, Майкл (2015). Нейробиология социального поведения: к пониманию просоциального и антисоциального мозга. Лондонг: Elsevier Science. п. 85.
- ^ Роллс, Эдмунд Т. (2006). «Глава 5 Нейрофизиология и функции орбитофронтальной коры». В Zald, David H .; Раух, Скотт Л. (ред.). Орбитофронтальная кора. Нью-Йорк: Издательство Оксфордского университета.
- ^ Охота LT; Малаласекера WMN; де Беркер АО; Миранда Б; Фермер S; Behrens TEJ; Kennerley SW (26 сентября 2018 г.). «Тройная диссоциация внимания и вычислений решений в префронтальной коре». Природа Неврология. 21 (9): 1471–1481. Дои:10.1038 / s41593-018-0239-5. ЧВК 6331040. PMID 30258238.
- ^ Шульц, Вольфрам; Тремблей, Леон (2006). «Глава 7: Участие орбитофронтальных нейронов приматов в вознаграждении, неопределенности и обучении 173 Вольфрам Шульц и Леон Трембле». В Zald, David H .; Раух, Скотт: Л. (ред.). Орбитофронтальная кора. Нью-Йорк: Издательство Оксфордского университета.
- ^ Шенбаум Г., Такахаши Ю., Лю Т., МакДанналд М. (2011). "Имеет ли значение сигнал орбитофронтальной коры?". Летопись Нью-Йоркской академии наук. 1239 (1): 87–99. Bibcode:2011НЯСА1239 ... 87С. Дои:10.1111 / j.1749-6632.2011.06210.x. ЧВК 3530400. PMID 22145878.
- ^ Берлин, HA; Rolls, ET; Иверсен, SD (октябрь 2005 г.). «Пограничное расстройство личности, импульсивность и орбитофронтальная кора». Архив клинической нейропсихологии. 20 (7): 862–863.
- ^ Ха, Сунджи; Сон, Ин-Юнг; Ким, Намвук; Сим, Хён Чжон; Cheon, Keun-Ah (декабрь 2015 г.). «Характеристики мозга при расстройствах аутистического спектра: структура, функции и возможности взаимодействия на протяжении всей жизни». Экспериментальная нейробиология. 24 (4): 273–284. Дои:10.5607 / en.2015.24.4.273. ISSN 1226-2560. ЧВК 4688328. PMID 26713076.
- ^ Jackowski, AP; Араужо Филью, GM; Almeida, AG; Araújo, CM; Рейс, М; Нери, Ф; Батиста, Ирландия; Сильва, I; Ласерда, Алабама (июнь 2012 г.). «Участие орбитофронтальной коры головного мозга в психических расстройствах: обновление результатов нейровизуализации». Revista Brasileira de Psiquiatria. 34 (2): 207–12. Дои:10.1590 / S1516-44462012000200014. PMID 22729418.
- ^ Милад, MR; Раух, С.Л. (декабрь 2007 г.). «Роль орбитофронтальной коры при тревожных расстройствах». Летопись Нью-Йоркской академии наук. 1121 (1): 546–61. Bibcode:2007НЯСА1121..546М. Дои:10.1196 / летопись.1401.006. PMID 17698998.
- ^ Накао, Т; Окада, К; Канба, С. (август 2014 г.). «Нейробиологическая модель обсессивно-компульсивного расстройства: данные последних нейропсихологических и нейровизуализационных данных». Психиатрия и клинические неврологии. 68 (8): 587–605. Дои:10.1111 / шт.12195. PMID 24762196.
- ^ Fineberg, NA; Potenza, MN; Чемберлен, SR; Берлин, HA; Menzies, L; Бечара, А; Саакян, Б.Дж.; Роббинс, TW; Bullmore, ET; Холландер, Э (февраль 2010 г.). «Исследование компульсивного и импульсивного поведения, от животных моделей до эндофенотипов: обзор повествования». Нейропсихофармакология. 35 (3): 591–604. Дои:10.1038 / npp.2009.185. ЧВК 3055606. PMID 19940844.
- ^ Милад, MR; Раух, С.Л. (январь 2012 г.). «Обсессивно-компульсивное расстройство: за пределами сегрегированных кортико-полосатых путей». Тенденции в когнитивных науках. 16 (1): 43–51. Дои:10.1016 / j.tics.2011.11.003. ЧВК 4955838. PMID 22138231.
- ^ Ваги, М; Роббинс, Т. "ЗАДАЧНЫЕ ФУНКЦИОНАЛЬНЫЕ НЕЙРОИМИЗАЦИОННЫЕ ИССЛЕДОВАНИЯ ОБСЕССИВНО-КОМПУЛЬСИВНОГО РАССТРОЙСТВА: ОБЗОР НА ГИПОТЕЗЕ". В Питтенгер, Кристофер (ред.). Обсессивно-компульсивное расстройство, феноменология, патофизиология и лечение. Издательство Оксфордского университета. С. 239–240.
- ^ Schoenbaum, G; Чанг, CY; Lucantonio, F; Такахаши, Ю.К. (декабрь 2016 г.). «Нестандартное мышление: орбитофронтальная кора, воображение и как мы можем лечить зависимость». Нейропсихофармакология. 41 (13): 2966–2976. Дои:10.1038 / npp.2016.147. ЧВК 5101562. PMID 27510424.
- ^ Кооб Г.Ф .; Волков, Н.Д. (январь 2010 г.). «Нейросхема зависимости». Нейропсихофармакология. 35 (1): 217–38. Дои:10.1038 / npp.2009.110. ЧВК 2805560. PMID 19710631.
- ^ Мурман, Делавэр (2 февраля 2018 г.). «Роль орбитофронтальной коры в употреблении алкоголя, злоупотреблении и зависимости». Прогресс в нейропсихофармакологии и биологической психиатрии. 87 (Pt A): 85–107. Дои:10.1016 / j.pnpbp.2018.01.010. ЧВК 6072631. PMID 29355587.
- ^ Gowin, JL; Макки, S; Паулюс, депутат (1 сентября 2013 г.). «Измененная обработка рисков у потребителей психоактивных веществ: дисбаланс между болью и выгодой». Наркотическая и алкогольная зависимость. 132 (1–2): 13–21. Дои:10.1016 / j.drugalcdep.2013.03.019. ЧВК 3748224. PMID 23623507.
Лица с SUD демонстрируют несколько аномалий обработки во время принятия решений о принятии риска, которые включают измененную оценку вариантов (VMPFC) и результатов (OFC и полосатое тело), плохую оценку неопределенности (ACC и островная кора), снижение исполнительного контроля (DLPFC), и ослабленное влияние эмоциональной значимости (миндалевидное тело) и снижение чувствительности к соматическим маркерам (соматосенсорная кора). Эти различия в нейронной обработке во время принятия риска среди людей с SUD были связаны с более низким поведением при выполнении задач, связанных с принятием риска, и более обширной историей употребления психоактивных веществ.
- ^ Чейз, HW; Айкхофф, SB; Laird, AR; Хогарт, Л. (15 октября 2011 г.). «Нейронная основа обработки лекарственного стимула и тяги: метаанализ оценки вероятности активации». Биологическая психиатрия. 70 (8): 785–93. Дои:10.1016 / j.biopsych.2011.05.025. ЧВК 4827617. PMID 21757184.
Медиальная область OFC показала большую активацию лекарственными сигналами по сравнению с контрольными сигналами и постоянно активировалась в подгруппе, не обращающейся за лечением. Есть веские доказательства того, что эта область играет роль в аппетитном поведении и принятии решений (86, 87), в частности, в отношении ожиданий вознаграждения (88), предсказываемых условными стимулами (89–94), которые могут контролировать выбор инструментальных действий
- ^ Lucantonio, F; Caprioli, D; Шенбаум, Г. (январь 2014 г.). «Переход от« модельного »к« безмодельному »поведенческому контролю при зависимости: вовлечение орбитофронтальной коры и дорсолатерального полосатого тела». Нейрофармакология. 76 Pt B: 407–15. Дои:10.1016 / j.neuropharm.2013.05.033. ЧВК 3809026. PMID 23752095.
- ^ Рубиа, К. (15 июня 2011 г.). ""Холодная «нижняя лобно-стриатальная дисфункция при синдроме дефицита внимания / гиперактивности по сравнению с« горячей »вентромедиальной орбитофронтально-лимбической дисфункцией при расстройстве поведения: обзор». Биологическая психиатрия. 69 (12): e69–87. Дои:10.1016 / j.biopsych.2010.09.023. PMID 21094938.
- ^ Rosell, DR; Сивер, LJ (июнь 2015 г.). «Нейробиология агрессии и насилия». Спектры ЦНС. 20 (3): 254–79. Дои:10.1017 / S109285291500019X. PMID 25936249.
- ^ а б Rolls, ET (сентябрь 2016 г.). "Теория депрессии, не связанная с вознаграждением" (PDF). Неврология и биоповеденческие обзоры. 68: 47–58. Дои:10.1016 / j.neubiorev.2016.05.007. PMID 27181908.
- ^ Дж. Уилсон; М. Дженкинсон; И. Э. Т. де Араужо; Мортен Л. Крингельбах; Э. Т. Роллс & Питер Джеззард (Октябрь 2002 г.). «Быстрая, полностью автоматизированная глобальная и локальная оптимизация магнитного поля для фМРТ человеческого мозга». NeuroImage. 17 (2): 967–976. Дои:10.1016 / S1053-8119 (02) 91172-9. PMID 12377170.
- ^ Ahmari, SE; Догерти, Д. Д. (август 2015 г.). «Рассечение цепей ОКД: от моделей на животных к целевому лечению». Депрессия и тревога. 32 (8): 550–62. Дои:10.1002 / da.22367. ЧВК 4515165. PMID 25952989.
- ^ Вертеки, Пьетро; Лоттем, Эран; Сарра, Дарио; Годиньо, Беатрис; Тревес, Исаак; Quendera, Tiago; Лохуис, Маттис Николай Ауд; Майнен, Закари Ф. (2020-04-08). «Решения, основанные на выводах в задаче по поиску пищи в скрытом состоянии: дифференциальные вклады префронтальных областей коры головного мозга». Нейрон. 106 (1): 166–176.e6. Дои:10.1016 / j.neuron.2020.01.017. ISSN 0896-6273. PMID 32048995.
- ^ Сноуден Дж. С .; Bathgate D .; Варма А .; Blackshaw A .; Гиббонс З. С .; Нири Д. (2001). «Отчетливые поведенческие профили при лобно-височной деменции и семантической деменции». J Neurol Neurosurg Psychiatry. 70 (3): 323–332. Дои:10.1136 / jnnp.70.3.323. ЧВК 1737271. PMID 11181853.
- ^ а б Paulus M. P .; Хозак Н.Е .; Заушер Б. Э .; Фрэнк Л .; Браун Г. Г .; Брафф Д. Л .; Шукит М. А. (2002). «Поведенческие и функциональные данные нейровизуализации для префронтальной дисфункции у субъектов, зависимых от метамфетамина». Нейропсихофармакология. 26 (1): 53–63. Дои:10.1016 / s0893-133x (01) 00334-7. PMID 11751032.
- ^ а б Топлак М. Э .; Jain U .; Таннок Р. (2005). «Исполнительные и мотивационные процессы у подростков с синдромом дефицита внимания и гиперактивности (СДВГ)». Поведенческие и мозговые функции. 1 (1): 8–20. Дои:10.1186/1744-9081-1-8. ЧВК 1183187. PMID 15982413.
- ^ а б Вердехо-Гарсия А .; Bechara A .; Рекнор Э. С .; Перес-Гарсия М. (2006). «Исполнительная дисфункция у лиц, зависимых от психоактивных веществ во время употребления наркотиков и воздержания: исследование поведенческих, когнитивных и эмоциональных коррелятов зависимости». Журнал Международного нейропсихологического общества. 12 (3): 405–415. Дои:10,1017 / с 1355617706060486. PMID 16903133.
- ^ Ча, Джиук; Гринберг, Цафрир; Карлсон, Джошуа М .; ДеДора, Дэниел Дж .; Хаджак, Грег; Мухика-Пароди, Лилианн Р. (12 марта 2014 г.). «Общесистемные структурные и функциональные показатели предсказывают генерализацию страха вентромедиальной префронтальной коры: последствия для генерализованного тревожного расстройства». Журнал неврологии. 34 (11): 4043–4053. Дои:10.1523 / JNEUROSCI.3372-13.2014. ISSN 0270-6474. ЧВК 6705282. PMID 24623781.
- ^ Волков Н.Д .; Фаулер Дж. (2000). «Зависимость - болезнь принуждения и влечения: поражение орбитофронтальной коры». Кора головного мозга. 10 (3): 318–325. Дои:10.1093 / cercor / 10.3.318. PMID 10731226.
- ^ Чемберлен С. Р .; Одлауг Б. Л .; Boulougouris V .; Файнберг Н. А .; Грант Дж. Э. (2009). «Трихотилломания: нейробиология и лечение». Неврология и биоповеденческие обзоры. 33 (6): 831–842. Дои:10.1016 / j.neubiorev.2009.02.002. PMID 19428495.
- ^ Мензис Л. (2008). «Объединение данных нейровизуализационных и нейропсихологических исследований обсессивно-компульсивного расстройства: пересмотр орбитофронто-полосатой модели». Неврология и биоповеденческие обзоры. 32 (3): 525–549. Дои:10.1016 / j.neubiorev.2007.09.005. ЧВК 2889493. PMID 18061263.
- ^ а б Текин С .; Каммингс Дж. Л. (2002). «Фронтально-подкорковые нейронные цепи и клиническая нейропсихиатрия: обновление». Журнал психосоматических исследований. 53 (2): 647–654. Дои:10.1016 / s0022-3999 (02) 00428-2. PMID 12169339.
- ^ Рахман С .; Саакян Б. Дж .; Hodges J. R .; Rogers R.D .; Роббинс Т. В. (1999). «Специфические когнитивные нарушения при раннем поведенческом варианте лобно-височной деменции». Мозг. 122 (8): 1469–1493. Дои:10.1093 / мозг / 122.8.1469. PMID 10430832.
- ^ Сили В. В .; Crawford R .; Расковский К .; Kramer J. H .; Weiner M .; Miller B.L .; Горно-Темпини Л. (2008). «Атрофия лобной паралимбической сети при очень легком поведенческом варианте лобно-височной деменции». Архив неврологии. 65 (2): 249–255. Дои:10.1001 / archneurol.2007.38. ЧВК 2544627. PMID 18268196.
- ^ Чибане, ИС; Буше, О; Dubeau, F; Тран, TPY; Мохамед, я; Маклахлан, Р. Сэдлер, РМ; Desbiens, R; Карман, L; Нгуен, Дания (ноябрь 2017 г.). «Орбитофронтальная эпилепсия: серия случаев и обзор литературы». Эпилепсия и поведение. 76: 32–38. Дои:10.1016 / j.yebeh.2017.08.038. PMID 28928072.
- ^ Золото, JA; Шер, Y; Мальдонадо, младший (2016). "Эпилепсия лобной доли: учебник для психиатров и систематический обзор психиатрических проявлений". Психосоматика. 57 (5): 445–64. Дои:10.1016 / j.psym.2016.05.005. PMID 27494984.
- ^ Смит, младший; Силлай, К; Винклер, П; King, DW; Лоринг, DW (2004). «Орбитофронтальная эпилепсия: электроклинический анализ хирургических случаев и обзор литературы». Стереотаксическая и функциональная нейрохирургия. 82 (1): 20–5. Дои:10.1159/000076656. PMID 15007215.
- ^ Роллс Э. Т .; Хорнак Дж .; Wade D .; МакГрат Дж. (1994). «Эмоциональное обучение у пациентов с социальными и эмоциональными изменениями, связанными с повреждением лобной доли». J Neurol Neurosurg Psychiatry. 57 (12): 1518–1524. Дои:10.1136 / jnnp.57.12.1518. ЧВК 1073235. PMID 7798983.
- ^ Bechara A .; Damasio A. R .; Damasio H .; Андерсон С.В. (1994). «Нечувствительность к будущим последствиям после повреждения префронтальной коры головного мозга человека». Познание. 50 (1–3): 7–15. Дои:10.1016/0010-0277(94)90018-3. PMID 8039375.
- ^ Стоун В.Е .; Барон-Коэн С .; Найт Р. Т. (1998a). «Вклад лобной доли в теорию разума». Журнал медицинских исследований. 10 (5): 640–656. CiteSeerX 10.1.1.330.1488. Дои:10.1162/089892998562942. PMID 9802997.