Примас - Primate
| Приматы | |
|---|---|
  | |
| Некоторые семейства приматов, сверху вниз: Daubentoniidae, Tarsiidae, Лемуриды, Lorisidae, Cebidae, Callitrichidae, Ателиды, Cercopithecidae, Hylobatidae, Гоминиды | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Млекопитающие |
| Зеркальный заказ: | Приматоморфа |
| Заказ: | Приматы Линней, 1758[1] |
| Подзаказы | |
сестра: Dermoptera | |
 | |
| Ареал нечеловеческих приматов (зеленый) | |
| Синонимы | |
Plesiadapiformes (кладистски включая приматов[2]) | |
А примат (/ˈпраɪмeɪт/ (![]() Слушать) PRY-майт ) (от латинского примат, из примус: "prime, first rank") является евтерский млекопитающее составляющий таксономический порядок Приматы (/праɪˈмeɪтяz/). Приматы возникли 85–55 миллионов лет назад сначала из малых земной млекопитающих, которые приспособились к жизни на деревьях тропические леса: многие характеристики приматов представляют собой приспособления к жизни в этой сложной среде, в том числе большой мозг, Острота зрения, цветовое зрение, а плечевой пояс позволяя большие движения в плечевом суставе и ловкие руки. Приматы различаются по размеру от Мышиный лемур мадам Берты, который весит 30 г (1 унцию), восточная горилла, весом более 200 кг (440 фунтов). Есть 190–448 разновидность живых приматов, в зависимости от используемой классификации. Продолжают открываться новые виды приматов: более 25 видов были описаны в первом десятилетии 2000-х годов и одиннадцать - с 2010 года.
Слушать) PRY-майт ) (от латинского примат, из примус: "prime, first rank") является евтерский млекопитающее составляющий таксономический порядок Приматы (/праɪˈмeɪтяz/). Приматы возникли 85–55 миллионов лет назад сначала из малых земной млекопитающих, которые приспособились к жизни на деревьях тропические леса: многие характеристики приматов представляют собой приспособления к жизни в этой сложной среде, в том числе большой мозг, Острота зрения, цветовое зрение, а плечевой пояс позволяя большие движения в плечевом суставе и ловкие руки. Приматы различаются по размеру от Мышиный лемур мадам Берты, который весит 30 г (1 унцию), восточная горилла, весом более 200 кг (440 фунтов). Есть 190–448 разновидность живых приматов, в зависимости от используемой классификации. Продолжают открываться новые виды приматов: более 25 видов были описаны в первом десятилетии 2000-х годов и одиннадцать - с 2010 года.
Приматы делятся на два разных подзаказы (см. диаграмму в разделе «История терминологии»). Первый подотряд называется Strepsirrhines (от греческого 'закрученный нос или закрученный нос'), который содержит лемуры, Galagos, и лорисиды. Этих приматов можно встретить по всей Африке, на Мадагаскаре, Индии и Юго-Восточной Азии. Разговорные названия видов, оканчивающиеся на носовой обратитесь к ринарий приматов. Второй подотряд называется haplorhines, который содержит «сухоносых» приматов (от греч. «простой нос») в долгопят, обезьяна, и обезьяна клады. В последнюю из этих групп входят люди. Обезьяны (инфразаказ называется Симиобразные от греческого слова simos, что означает «плосконосый») относятся к обезьянам и обезьянам, которые могут быть классифицированы как обезьяны Старого Света и обезьяны под инфраотрядом катар (от греческого «узконосый») или как обезьяны Нового Света под отрядом Platyrrhines (от греч. «плосконосый»). Сорок миллионов лет назад обезьяны из Африки мигрировали в Южную Америку. дрейфует на обломках (предположительно), которые дали начало пяти семействам обезьян Нового Света. Остальные обезьяны (catarrhines) разделились на обезьян (Гоминоидея ) и обезьяны Старого Света (Cercopithecoidea ) примерно двадцать пять миллионов лет назад. Общие виды обезьян включают (Старый мир ) бабуины, макаки, гиббоны, и большие обезьяны; и (Новый мир ) капуцины, вопли и беличьи обезьяны.
У приматов большой мозг (по сравнению с размером тела) по сравнению с другими млекопитающими, а также повышенная зависимость от остроты зрения за счет чувства зрения. запах, которая является доминирующей сенсорной системой у большинства млекопитающих. Эти черты более развиты у обезьян и приматов и заметно меньше у лорисов и лемуров. Некоторые приматы трихроматы, с тремя независимыми каналами для передачи цветовой информации. Кроме обезьяны (включая люди ), у приматов есть хвосты. У большинства приматов также есть противоборствующие пальцы. Многие виды сексуально диморфный; различия могут включать мышца масса, толстый распределение, тазовый ширина, размер клыков, распределение волос и окраска. Приматы развиваются медленнее, чем другие млекопитающие того же размера, позже достигают зрелости и имеют более длительную продолжительность жизни. В зависимости от вида взрослые особи могут жить в одиночестве, в спариваемых парах или в группах до сотен членов. Некоторые приматы, в том числе гориллы, люди, и бабуины, прежде всего земной скорее, чем древесный, но у всех видов есть приспособления для лазания по деревьям. Древесный движение используемые техники включают прыжки с дерева на дерево и раскачивание между ветвями деревьев (брахиация ); техники наземного передвижения включают ходьбу на двух конечностях (двуногие ) и модифицированная ходьба на четырех конечностях (ходьба на костяшках пальцев ).
Приматы - одни из самых социальных животных, образующие пары или семейные группы, гаремы с одним самцом и группы, состоящие из нескольких самцов и самок. Приматы, кроме человека, имеют по крайней мере четыре типа социальные системы, многие из которых определяются количеством движений девушек-подростков между группами. Большинство видов приматов по крайней мере частично остаются древесными: исключение составляют люди, некоторые другие человекообразные обезьяны и бабуины, которые оставили деревья на землю и теперь населяют все континенты.
Тесное взаимодействие между людьми и приматами (НЧП) может создавать возможности для передачи зоонозы, особенно вирусные заболевания, в том числе герпес, корь, Эбола, бешенство, и гепатит. Тысячи нечеловеческих приматов используются в исследованиях по всему миру из-за их психологического и физиологического сходства с людьми. Около 60% видов приматов находятся под угрозой исчезновения. Общие угрозы включают вырубка леса, фрагментация леса, обезьяна едет и охота на приматов для использования в медицине, в качестве домашних животных и в пищу. Крупномасштабная вырубка тропических лесов для сельскохозяйственных нужд больше всего угрожает приматам.
История терминологии
Английское название «приматы» происходит от Старофранцузский или же Французский примат, от латинского слова примат, из примус («премьер, первый разряд»).[3] Имя было дано Карл Линней потому что он считал это «высшим» порядком животных.[4] Отношения между различными группами приматов не были четко поняты до относительно недавнего времени, поэтому часто используемые термины несколько сбиты с толку. Например, "обезьяна "использовался как альтернатива" обезьяны "или для любого бесхвостого, относительно человек -подобный примат.[5]
сэр Уилфрид Ле Гро Кларк был одним из приматологи который развил идею тенденций в эволюции приматов и методологию организации живых членов отряда в «восходящую серию», ведущую к людям.[6] Часто используемые названия групп приматов, такие как "Prosimians ", "обезьяны ", "меньшие обезьяны ", и "большие обезьяны "отражают эту методологию. Согласно нашему нынешнему пониманию эволюционной истории приматов, некоторые из этих групп являются парафилетический: парафилетическая группа - это группа, которая нет включают всех потомков общего предка группы.[7]
В отличие от методологии Кларка, современные классификации обычно идентифицируют (или называют) только те группы, которые монофилетический; то есть такая именованная группа включает все потомки общего предка группы.[8]
В кладограмма ниже показана одна из возможных классификационных последовательностей живых приматов:[9][10] Справа показаны группы, использующие общие (традиционные) имена.
| Prosimians обезьяны большие обезьяны люди меньшие обезьяны |




Все группы с научными названиями монофилетичны (т. Е. клады ), а последовательность научной классификации отражает эволюционную историю родственных линий. Группы с традиционными названиями показаны справа; они образуют «восходящую серию» (по Кларку, см. выше), и несколько групп являются парафилетическими:
- «прозимианы» содержат две монофилетические группы (подотряд Strepsirrhini, или лемуры, лори и союзники, а также долгопяты подотряда Haplorhini); это парафилетическая группа, потому что она исключает Simiiformes, которые также являются потомками общих предков приматов.
- «обезьяны» включают две монофилетические группы, обезьяны Нового Света и обезьяны Старого Света, но являются парафилетическими, поскольку исключают гоминоидов, надсемейство Hominoidea, также потомков общего предка Simiiformes.
- "обезьяны "в целом, и"большие обезьяны ", являются парафилетическими, если термины используются так, что исключают люди.
Таким образом, члены двух наборов групп и, следовательно, имена не совпадают, что вызывает проблемы при связывании научных названий с общими (обычно традиционными) названиями. Рассмотрим надсемейство Hominoidea: с точки зрения общих имен справа, эта группа состоит из обезьян и людей, и не существует единого общего названия для всех членов группы. Одно из решений - создать новое общее название, в данном случае «гуманоиды». Другая возможность - расширить использование одного из традиционных имен. Например, в его книге 2005 г. позвоночное животное палеонтолог Бентон писал: «К обезьянам, Hominoidea, сегодня относятся гиббоны и орангутанги ... гориллы и шимпанзе ... и люди»;[11] таким образом, Бентон использовал слово «обезьяны» для обозначения «гоминоидов». В этом случае группа, ранее называемая «обезьянами», теперь должна быть идентифицирована как «нечеловеческие обезьяны».
По состоянию на 2005 г.[Обновить], нет единого мнения относительно того, следует ли принимать традиционные (то есть общие), но парафилетические имена или использовать только монофилетические имена; или использовать «новые» общеупотребительные имена или адаптации старых. Оба конкурирующих подхода можно найти в биологических источниках, часто в одной и той же работе, а иногда и одним и тем же автором. Таким образом, Бентон определяет «обезьяны», чтобы включить людей, затем он неоднократно использует «обезьяноподобный», чтобы означать «как обезьяна, а не человек»; и при обсуждении реакции других людей на новую окаменелость он пишет о «утверждениях, что Оррорин ... был скорее обезьяной, чем человеком ".[12]
Классификация живых приматов
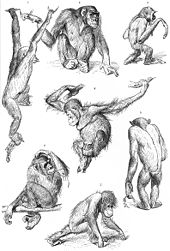


Список семейств ныне живущих приматов приведен ниже вместе с одной из возможных классификаций по рангам между отрядом и семейством.[1][9][13][14] Также используются другие классификации. Например, альтернативная классификация живых Strepsirrhini делит их на два инфраотряда, Lemuriformes и Lorisiformes.[15]
- Приматы отряда
- Подотряд Strepsirrhini: лемуры, галаго и лорисиды
- Инфразаказ Лемурообразные[а]
- Надсемейство Lemuroidea
- Семья Cheirogaleidae: карликовые лемуры и мыши-лемуры (34 вида)
- Семья Daubentoniidae: aye-aye (один вид)
- Семья Лемуриды: кольчатый лемур и союзники (21 вид)
- Семья Lepilemuridae: спортивные лемуры (26 видов)
- Семья Индрииды: шерстистые лемуры и их союзники (19 видов)
- Надсемейство Lorisoidea
- Надсемейство Lemuroidea
- Инфразаказ Лемурообразные[а]
- Подотряд Haplorhini: долгопяты, обезьяны и обезьяны
- Инфразаказ Tarsiiformes
- Семья Tarsiidae: долгопяты (11 видов)
- Инфразаказ Симиобразные (или Anthropoidea)
- Парвордер Платиррини: Обезьяны Нового Света
- Семья Callitrichidae: мартышки и тамарины (42 вида)
- Семья Cebidae: капуцины и беличьи обезьяны (14 видов)
- Семья Aotidae: ночные или совообразные обезьяны (douroucoulis) (11 видов)
- Семья Pitheciidae: titis, sakis и uakaris (43 вида)
- Семья Ателиды: ревун, паук, шерстистый паук и шерстистые обезьяны (29 видов)
- Парвордер Катаррини
- Надсемейство Cercopithecoidea
- Семья Cercopithecidae: Обезьяны Старого Света (138 видов)
- Надсемейство Гоминоидея
- Семья Hylobatidae: гиббоны или «малые обезьяны» (18 видов)
- Семья Гоминиды: человекообразные обезьяны, включая человека (8 видов)
- Надсемейство Cercopithecoidea
- Парвордер Платиррини: Обезьяны Нового Света
- Инфразаказ Tarsiiformes
- Подотряд Strepsirrhini: лемуры, галаго и лорисиды
Орден Предстоятелей был учрежден Карл Линней в 1758 г., в десятое издание его книги Systema Naturae,[18] для родов Гомо (люди ), Симиа (Другой обезьяны и обезьяны), Лемур (просимиане) и Веспертилио (летучие мыши). В первом издании той же книги (1735 г.) он использовал имя Антропоморфные за Гомо, Симиа и Bradypus (ленивцы).[19] В 1839 г. Анри Мари Дюкротэ де Бленвиль вслед за Линнеем и подражая его номенклатуре, установил порядки Секундаты (включая подзаказы Рукокрылые, Насекомоядные и Хищник ), Tertiates (или же Glires ) и Четвертичные (включая Gravigrada, Пахидерматы и Руминантия ),[20] но эти новые таксоны не были приняты.
Прежде чем Андерсон и Джонс представили классификацию Strepsirrhini и Haplorhini в 1984 году,[21] (за которой следует работа Маккенны и Белла 1997 г. Классификация млекопитающих: выше уровня вида ),[22] Приматы разделились на два надсемейства: Просимы и Антропоидея.[23] Просимии включали все Prosimians: Strepsirrhini плюс долгопяты. Anthropoidea содержала все обезьяны.
Филогения и генетика
|
Заказ Приматы - часть клады Euarchontoglires, который вложен в кладу Евтерия класса Млекопитающие. Недавние молекулярно-генетические исследования приматов, Colugos, и деревья показал, что два вида колуго более тесно связаны с приматами, чем с землероками,[24] хотя когда-то землеройки считались приматами.[25] Эти три заказа составляют клады Euarchonta. Сочетание клады с кладой Glires (состоит из Rodentia и Зайцеобразные ) образует кладу Euarchontoglires. По-разному, как Euarchonta, так и Euarchontoglires считаются надотрядами. Некоторые ученые считают Dermoptera подотрядом приматов и используют подотряд Euprimates для «настоящих» приматов.[26]
Эволюция
Считается, что происхождение приматов восходит к Граница мела и палеогена или около 63–74 (моя ),[27][28][29][30][31] даже несмотря на то, что самые старые известные приматы от летописи окаменелостей до позднего палеоцена Африки, около 57 млн лет назад (Altiatlasius )[32] или переход палеоцена-эоцена на северных континентах, c. 55 млн лет назад (Кантий, Donrussellia, Алтаний, Плезиадапис и Тейльхардина ).[33][34][35] Другие исследования, в том числе исследования молекулярных часов, оценили происхождение ветви приматов в середине мелового периода, около 85 млн лет назад.[36][37][38]
По современным кладистический По подсчетам, Приматы монофилетический. Подотряд Strepsirrhini, "мокрый "приматы, как принято считать, отделились от примитивной линии примитивов около 63 млн лет назад,[39] хотя более ранние даты также поддерживаются.[40] Семь семейств Strepsirrhine - это пять родственных лемур семьи и две оставшиеся семьи, которые включают лорисиды и Galagos.[1][13] Перенос старых схем классификации Lepilemuridae в Лемуриды и Galagidae в Lorisidae, давая семейное распределение четыре-один вместо пяти-двух, как представлено здесь.[1] Вовремя эоцен, на большей части северных континентов преобладали две группы: Adapiforms и омомииды.[41][42] Первые считаются членами Strepsirrhini, но не имеют частый гребень как современные лемуры; недавний анализ показал, что Darwinius masillae вписывается в эту группировку.[43] Последние были тесно связаны с долгопятами, обезьянами и обезьянами. Как эти две группы относятся к дошедшим до наших дней приматам, неясно. Омомииды погибли около 30 млн лет назад,[42] в то время как адапиформы выжили примерно до 10 млн лет назад.[44]
Согласно генетическим исследованиям, лемуры Мадагаскара отклонились от лорисоидов примерно на 75 млн лет назад.[40] Эти исследования, а также хромосомные и молекулярные данные также показывают, что лемуры более тесно связаны друг с другом, чем с другими стрепсеринскими приматами.[40][45] Однако Мадагаскар отделился от Африки на 160 млн лет назад и от Индии на 90 млн лет назад.[46] Чтобы объяснить эти факты, предполагается, что основная популяция лемуров, состоящая из нескольких особей, достигла Мадагаскара из Африки через один рафтинг от 50 до 80 млн лет назад.[40][45][46] Были предложены другие варианты колонизации, такие как множественные колонизации из Африки и Индии,[41] но ни одно из них не подтверждено генетическими и молекулярными доказательствами.[40]

До недавнего времени да-да было трудно поместить в Strepsirrhini.[1] Были предложены теории, согласно которым его семейство, Daubentoniidae, было либо лемурообразным приматом (то есть его предки отделились от линии лемуров позже, чем лемуры и лорисы), либо сестринской группой по отношению ко всем остальным стрепсирринам. В 2008 году было подтверждено, что семья ай-айе наиболее тесно связана с другими малагасийскими лемурами, вероятно, происходящими от той же предковой популяции, которая колонизировала остров.[40]
Подотряд Haplorhini, приматы с простым носом или «сухоносые», состоят из двух сестринских клад.[1] Просимиан долгопяты в семействе Tarsiidae (монотипные в собственном инфраотряде Tarsiiformes) представляют собой наиболее базальный дивизии, возникшей около 58 млн лет назад.[47][48] Самый ранний из известных скелет гаплорин, старый долгопят 55 млн лет. Archicebus, был найден в центральном Китае,[49] подтверждая уже предполагаемое азиатское происхождение группы.[50] Инфразаказ Симиобразные (обезьяньи приматы, состоящие из обезьян и обезьян) появились около 40 млн лет назад,[42] возможно также в Азии; если так, то они рассредоточенный через Море Тетис вскоре после этого из Азии в Африку.[51] Есть две обезьяньи клады, обе парвордеры: Катаррини, который развился в Африке, состоящий из Обезьяны Старого Света, люди и другие обезьяны, и Platyrrhini, который развился в Южной Америке, состоящий из Обезьяны Нового Света.[1] Третья клада, которая включала эозимииды, возникла в Азии, но вымерла миллионы лет назад.[52]
Как и в случае с лемурами, происхождение обезьян Нового Света неясно. Молекулярные исследования конкатенированных ядерных последовательностей дали широко варьирующуюся предполагаемую дату расхождения между платирринами и катарринами, в пределах от 33 до 70 миллионов лет назад, в то время как исследования, основанные на митохондриальных последовательностях, дают более узкий диапазон от 35 до 43 миллионов лет назад.[34][53] Антропоидные приматы, возможно, пересекли Атлантический океан от Африки до Южной Америки во время эоцен к прыжки по островам при содействии Хребты Атлантического океана и пониженный уровень моря.[41] В качестве альтернативы одиночный рафтинг может объяснить эту трансокеанскую колонизацию. Из-за Континентальный дрифт В то время Атлантический океан не был таким широким, как сегодня.[41] Исследования показывают, что небольшой примат весом 1 кг (2,2 фунта) мог прожить 13 дней на плоту из растений.[54] Учитывая предполагаемую скорость течения и скорость ветра, это давало бы достаточно времени, чтобы совершить путешествие между континентами.

Обезьяны и обезьяны распространились из Африки в Европу и Азию, начиная с Миоцен.[55] Вскоре лори и долгопяты отправились в то же путешествие. Первые окаменелости гомининов были обнаружены в Северной Африке и датируются 5–8 миллионами лет назад.[42] Обезьяны Старого Света исчезли из Европы около 1,8 млн лет назад.[56] Молекулярные исследования и исследования окаменелостей обычно показывают, что современные люди произошли в Африке 100 000–200 000 лет назад.[57]
Хотя приматы хорошо изучены по сравнению с другими группами животных, несколько новых видов получили был обнаружен недавно, а генетические тесты выявили ранее неизвестные виды в известных популяциях. Таксономия приматов в 2001 г. перечислено около 350 видов приматов;[10] Автор, Колин Гровс, увеличил это число до 376 за свой вклад в третье издание Виды млекопитающих мира (MSW3).[1] Однако публикации с момента составления таксономии в MSW3 в 2003 г. увеличили число до 424 видов, или 658, включая подвиды.[14]
Гибриды
Примас гибриды обычно возникают в неволе,[58] но были примеры и в дикой природе.[59][60] Гибридизация происходит, когда ареал двух видов перекрывается с образованием гибридные зоны; гибриды могут быть созданы людьми, когда животных помещают в зоопарки, или из-за давления окружающей среды, такого как хищничество.[59] Межродовые гибридизации, гибриды разных родов также были обнаружены в дикой природе. Хотя они принадлежат к родам, которые существовали в течение нескольких миллионов лет, скрещивание между ними все еще происходит. Gelada и гамадрилья бабуин.[61]
Клоны
24 января 2018 года ученые Китая сообщили в журнале Клетка создание двух крабоядная макака клоны, названный Чжун Чжун и Хуа Хуа, с использованием сложный метод переноса ДНК это произвело Долли овца, в первый раз.[62][63][64][65][66]
Анатомия и психология
Голова

Череп приматов имеет большой куполообразный череп, что особенно заметно в антропоиды. Череп защищает большой мозг, что является отличительной чертой этой группы.[67] Эндокраниальный объем (объем внутри черепа) в три раза больше в люди чем у величайших нечеловеческих приматов, что отражает больший размер мозга.[68] Средний эндокраниальный объем составляет 1201 кубический сантиметр у человека, 469 см.3 в гориллы, 400 см3 в шимпанзе и 397 см3 в орангутаны.[68] Первичная эволюционная тенденция приматов заключалась в развитии мозга, в частности неокортекс (часть кора головного мозга ), который связан с чувственное восприятие, поколение моторные команды, пространственное мышление, сознательная мысль а у людей язык.[69] В то время как другие млекопитающие в значительной степени полагаются на свои Чувство обоняния, древесная жизнь приматов привела к тактильный, визуально доминирующая сенсорная система,[69] сокращение обонятельной области мозга и усложнение социального поведения.[70]
Глаза приматов обращены вперед на передней части черепа; бинокулярное зрение позволяет точно воспринимать расстояние, полезно для брахи предки всех человекообразных обезьян.[67] А костный гребень над глазницами укрепляет более слабые кости лица, которые подвергаются нагрузке во время жевания. Strepsirrhines есть заглазничная перемычка, кость вокруг глазницы, чтобы защитить глаза; напротив, высшие приматы, haplorhines, появились полностью закрытые розетки.[71]

Приматы демонстрируют эволюционную тенденцию к уменьшению морда.[72] Технически обезьяны Старого Света отличаются от обезьян Нового Света строением носа, а от обезьян - носом. расположение их зубов.[70] У обезьян Нового Света ноздри обращены вбок; у обезьян Старого Света они обращены лицом вниз.[70] Зубной паттерн приматов значительно различается; хотя некоторые потеряли большую часть своих резцы, все сохраняют хотя бы один нижний резец.[70] У большинства стрепсерин нижние резцы образуют частый гребень, который используется для ухода за шерстью и иногда для кормления.[70][73] У обезьян Старого Света восемь премоляры, по сравнению с 12 обезьянами Нового Света. Виды Старого Света делятся на обезьян и обезьян в зависимости от количества куспиды на их коренные зубы: у обезьян четыре, у обезьян пять[70] - хотя у людей их может быть четыре или пять.[74] Главный бугор коренного зуба гоминида (лицемер ) возник в ранней истории приматов, тогда как острие соответствующего примитивного нижнего коренного зуба (параконида) было потеряно. Просимианы отличаются неподвижностью верхней губы, влажным кончиком носа и обращенными вперед нижними передними зубами.
Тело

Приматы обычно имеют по пять пальцев на каждой конечности (пентадактилия ), с характерным типом кератина ноготь на концах каждого пальца руки и ноги. Нижние стороны рук и ног имеют чувствительные подушечки на кончики пальцев. Большинство из них противоборствующие пальцы, характерная черта приматов, наиболее развитая у людей, но не ограниченная этим отрядом, (опоссумы и коалы, например, тоже есть).[67] Большие пальцы позволяют некоторым видам использовать инструменты. У приматов комбинация противоположных больших пальцев, коротких ногтей (а не когтей) и длинных, закрывающихся внутрь пальцев является реликт наследственной практики захвата ветвей, что частично позволило некоторым видам развиться брахиация (качание руками от ветки к ветке дерева) как важное средство передвижения. На втором пальце каждой стопы у просимианов есть когтистые когти, называемые туалетные когти, которые они используют для ухода.[67]
Примат ключица является важным элементом грудной пояс; это позволяет плечевой сустав широкая мобильность.[72] По сравнению с обезьянами Старого Света, обезьяны имеют более подвижные плечевые суставы и руки из-за дорсального положения лопатка широкие грудные клетки, более плоские спереди назад, более короткий и менее подвижный позвоночник и с более низким позвонки значительно уменьшено - что приводит к потере хвоста у некоторых видов. Цепкие хвосты находятся в ателиды, в том числе ревун, паук, шерстистый паук, шерстистые обезьяны; И в капуцины.[75][76] Приматы-самцы имеют висячий пенис и мошоночные яички.[77][73]
Половой диморфизм

Половой диморфизм часто выставляется в обезьяны, хотя в большей степени у видов Старого Света (обезьяны и некоторые обезьяны), чем у видов Нового Света. Недавние исследования включают сравнение ДНК для изучения вариаций в выражении диморфизма у приматов и основных причин полового диморфизма. Приматы обычно имеют диморфизм в масса тела[78][79][80] и размер клыков[81][82][83] вместе с шерсть и цвет кожи.[84] Диморфизм может быть объяснен различными факторами, в том числе: система спаривания,[78][85] размер,[85] среда обитания и диета.[86]
Сравнительный анализ позволил лучше понять взаимосвязь между половой отбор, естественный отбор, и системы спаривания у приматов. Исследования показали, что диморфизм - это результат изменений как мужских, так и женских черт.[78][87] Онтогенетическое масштабирование, при котором происходит относительное расширение общей траектории роста, может дать некоторое представление о взаимосвязи между половым диморфизмом и моделями роста.[88] Некоторые свидетельства из летописи окаменелостей предполагают, что конвергентная эволюция диморфизма, и некоторые вымершие гоминиды вероятно, имел больший диморфизм, чем любой живой примат.[87]
Передвижение

Виды приматов движутся мимо брахиация, двуногие, прыгающий, древесные и наземные четвероногие, альпинизм, ходьба на костяшках пальцев или комбинацией этих методов. Некоторые прозимианы в основном вертикальные цепляющиеся и прыгуны. К ним относятся многие кустарники, все индрииды (т.е. сифакас, авахис и Индрис ), спортивные лемуры, и все долгопяты.[89] Другие прозимианы - древесные четвероногие и альпинисты. Некоторые из них также являются наземными четвероногими, а некоторые прыгуны. Большинство обезьян являются как древесными, так и наземными четвероногими и альпинистами. Гиббонс, Muriquis и паучьи обезьяны все брахиаты широко,[56] причем гиббоны иногда делают это в удивительно акробатической манере. Шерстистые обезьяны также иногда брахиаты.[90] Орангутаны используют похожую форму передвижения, называемую квадраманальным лазанием, при которой они используют свои руки и ноги, чтобы нести свое тяжелое тело через деревья.[56] Шимпанзе и гориллы костяшки пальцев[56] и может передвигаться на короткие расстояния на двух ногах. Хотя многочисленные виды, такие как австралопитеки и ранние гоминиды, продемонстрировали полностью двуногое передвижение, люди являются единственным существующим видом с этой чертой.[91]
Зрение

В эволюция цветового зрения у приматов уникален среди большинства евтерский млекопитающие. Пока пульт позвоночное животное предки приматов обладали трехцветное зрение (трихроматизм), ночной образ жизни, теплокровный, предки млекопитающих потеряли одну из трех колбочек в сетчатка вовремя Мезозойский эпоха. Рыбы, рептилии и птицы поэтому трехцветные или тетрахроматический, в то время как все млекопитающие, за исключением некоторых приматов и сумчатые,[92] дихроматы или монохроматы (полностью дальтоник).[73] Ночные приматы, такие как ночные обезьяны и Буш младенцы, часто бывают одноцветными. Катаррин обычно бывает трехцветным из-за дупликация гена красно-зеленых опсин ген, лежавший в основе их происхождения 30-40 миллионов лет назад.[73][93] Platyrrhines, с другой стороны, только в некоторых случаях трехцветный.[94] В частности, отдельные самки должны быть гетерозиготный для двух аллели гена опсина (красный и зеленый), расположенного на одном локус из Х хромосома.[73] Следовательно, самцы могут быть только двухцветными, а женщины - двухцветными или трехцветными. Цветовое зрение у Strepsirrhines не так хорошо изучено; тем не менее, исследования указывают на диапазон цветового зрения, аналогичный тому, который обнаруживается у плоских черепах.[73]
Как и катаррины, обезьяны-ревуны (семейство платирринов) демонстрируют обычный трихроматизм, который был прослежен до эволюционно недавнего времени. дупликация гена.[95] Обезьяны-ревуны - одни из самых специализированных листоедов обезьян Нового Света; фрукты не являются основной частью их рациона,[90] а тип листьев, которые они предпочитают потреблять (молодые, питательные и легкоусвояемые), можно определить только по красно-зеленому сигналу. Полевые исследования диетических предпочтений ревунов показывают, что рутинный трихроматизм был выбран окружающей средой.[94]
Поведение
Социальные системы
Ричард Рэнгэм заявил, что социальные системы приматов лучше всего классифицировать по количеству движений самок между группами.[96] Он предложил четыре категории:
- Системы женского переноса - самки уходят из группы, в которой родились. Женщины группы не будут тесно связаны между собой, тогда как мужчины останутся со своими натальными группами, и эта тесная связь может иметь влияние на социальное поведение. Формируемые группы обычно довольно небольшие.[96] Эту организацию можно увидеть в шимпанзе, где мужчины, которые обычно являются родственниками, будут сотрудничать в защите территории группы.[97] Среди обезьян Нового Света, паучьи обезьяны и Muriquis используйте эту систему.[98]

- Системы передачи мужчин - пока женщины остаются в своих родных группах, мужчины эмигрируют в подростковом возрасте. Полигинный и общества, состоящие из нескольких мужчин, относятся к этой категории. Размеры групп обычно больше.[96] Эта система распространена среди кошачий лемур, обезьяны капуцины и церкопитеки обезьяны.[56]
- Моногамный вид - связь самца и самки, иногда сопровождаемая малолетним потомством. Есть общая ответственность за родительскую опеку и территориальную защиту. Потомство покидает территорию родителей в подростковом возрасте.[96] Гиббонс по существу используют эту систему, хотя «моногамия» в этом контексте не обязательно означает абсолютную сексуальную верность.[99] Эти виды не живут большими группами.
- Одиночные виды - часто самцы, защищающие территории, на которых обитают несколько самок.[96] Этот тип организации встречается у таких прозиманов, как медленный лорис.[100] Орангутаны не защищают свою территорию, но эффективно имеют эту организацию.[101]
Известно, что встречаются и другие системы. Например, с обезьяны-ревуны и гориллы как самцы, так и самки обычно переходят из своей натальной группы по достижении половой зрелости, в результате чего образуются группы, в которых ни самцы, ни самки обычно не связаны.[90][102] Некоторые просимиане, колобин обезьяны и каллитрихид обезьяны также используют эту систему.[56]
Переход самок или самцов из их родной группы, вероятно, является адаптацией для избежания инбридинга.[103] Анализ записей о размножении колоний содержащихся в неволе приматов, представляющих множество различных видов, показывает, что младенческая смертность инбредного молодняка обычно выше, чем у неинбредного молодняка.[103][104] Этот эффект инбридинга на младенческую смертность, вероятно, в значительной степени является результатом повышенной экспрессии вредных рецессивных аллелей (см. Инбридинговая депрессия ).

Приматолог Джейн Гудолл, которые учились в Национальный парк Гомбе-Стрим, отметил общества ядерного синтеза у шимпанзе.[105] Есть деление когда основная группа в течение дня разделяется на корм, то слияние когда группа возвращается ночью, чтобы спать как группа. Эту социальную структуру можно также наблюдать в гамадрилья бабуин,[106] паучьи обезьяны[90] и бонобо.[106] В Gelada имеет аналогичную социальную структуру, в которой множество более мелких групп собираются вместе, чтобы сформировать временные стада до 600 обезьян.[106] Люди также образуют общества ядерного синтеза. В обществах охотников-собирателей люди образуют группы, состоящие из нескольких особей, которые могут разделяться для получения различных ресурсов.[107]
На эти социальные системы влияют три основных экологических фактора: распределение ресурсов, размер группы, и хищничество.[108] Внутри социальной группы существует баланс между сотрудничеством и конкуренцией. Кооперативное поведение у многих видов приматов включает: социальный уход (удаление кожные паразиты и очистка ран), совместное использование пищи и коллективная защита от хищников или территории. Агрессивное поведение часто сигнализирует о конкуренции за еду, места для сна или партнеров. Агрессия также используется при установлении иерархии доминирования.[108][109]
Межвидовые ассоциации
Известно, что несколько видов приматов объединяются в дикой природе. Некоторые из этих ассоциаций были тщательно изучены. в Тайский лес в Африке несколько видов координируют поведение против хищников. К ним относятся Обезьяна диана, Обезьяна Кэмпбелла мона, малая косоглазая обезьяна, западный красный колобус, король колобус (западный черно-белый колобус ), и закопченный мангабей, которые координируют вызовы тревоги против хищников.[110] Среди хищников этих обезьян есть обыкновенный шимпанзе.[111]
В краснохвостая обезьяна ассоциируется с несколькими видами, в том числе с западным красным колобусом, синяя обезьяна, Волчья обезьяна мона, мантии гереза, черный хохлатый мангабей и Болотная обезьяна Аллена.[106] На некоторые из этих видов охотятся обычные шимпанзе.[112]
В Южной Америке, беличьи обезьяны ассоциироваться с обезьяны капуцины.[113] Это может быть больше связано с преимуществами кормления беличьих обезьян, чем с преимуществами борьбы с хищниками.[113]
Коммуникация

Лемуры, лори, долгопяты, а обезьяны Нового Света полагаются на обонятельные сигналы по многим аспектам социального и репродуктивного поведения.[69] Специализированные сальники используются для отмечать территории с феромоны, которые обнаруживаются вомероназальный орган; этот процесс составляет большую часть коммуникативного поведения этих приматов.[69] У обезьян Старого Света эта способность в основном рудиментарный, регрессировав как трехцветный глаза эволюционировали, чтобы стать основным органом чувств.[114] Приматы также используют вокализацию, жесты и мимику для передачи психологического состояния.[115][116] Лицевая мускулатура очень развита у приматов, особенно у обезьян и обезьян, что делает возможным сложное общение на лице. Как и люди, шимпанзе могут различать лица знакомых и незнакомых людей.[117] Жесты рук и кистей также являются важными формами общения для человекообразных обезьян, и один жест может иметь несколько функций.[116]
В Филиппинский долгопят, имеет высокочастотный предел слуховой чувствительности приблизительно 91 кГц с доминирующей частотой 70 кГц. Такие значения являются одними из самых высоких среди всех наземных млекопитающих и являются относительно экстремальным примером ультразвуковой связи. Для филиппинских долгопятов ультразвуковые вокализации могут представлять собой частный канал связи, который препятствует обнаружению хищниками, добычей и конкурентами, повышает энергетическую эффективность или улучшает обнаружение на фоне низкочастотного фонового шума.[118] Мужской обезьяны-ревуны являются одними из самых громких наземных млекопитающих, и их рев можно услышать на расстоянии до 4,8 км (3,0 мили).[119] Рев производит модифицированный гортань и увеличенный подъязычная кость который содержит воздушный мешок.[120] Считается, что эти призывы относятся к межгрупповому расстоянию и территориальной защите, а также, возможно, к охране партнера.[121] В верветка обезьяна подает отчетливый сигнал тревоги для каждого из по крайней мере четырех разных хищников, а реакция других обезьян варьируется в зависимости от этого сигнала. Например, если сигнал тревоги сигнализирует питону, обезьяны забираются на деревья, тогда как сигнал тревоги орла заставляет обезьян искать укрытие на земле.[122] У многих нечеловеческих приматов анатомия голоса позволяет воспроизводить человеческую речь, но у них отсутствует надлежащая мозговая проводка.[123] Подобные гласные звуки голоса были записаны у бабуинов, что имеет значение для происхождения речи у людей.[124]
Временной диапазон эволюции человек язык и / или его анатомические предпосылки происходят, по крайней мере в принципе, от филогенетического расхождения Гомо (От 2,3 до 2,4 миллиона лет назад) Сковорода (5-6 миллионов лет назад) до появления полного поведенческая современность около 50 000–150 000 лет назад. Мало кто спорит, что Австралопитек вероятно, не хватало вокального общения, значительно более сложного, чем у большие обезьяны в целом.[125]
История жизни

Приматы развиваются медленнее, чем другие млекопитающие.[56] Все младенцы приматов на грудном вскармливании их матерями (за исключением некоторых человеческих культур и различных приматов, выращиваемых в зоопарках), и полагаются на них в уходе и транспортировке.[56] У некоторых видов младенцев защищают и переносят самцы в группе, особенно самцы, которые могут быть их отцами.[56] Другие родственники младенца, такие как братья, сестры и тети, также могут участвовать в уходе за ним.[56] Большинство матерей-приматов перестают овуляция при грудном вскармливании младенца; как только младенец отлученный от груди мать снова может воспроизводить потомство.[56] Это часто приводит к конфликту при отлучении от грудного ребенка, который пытается продолжить грудное вскармливание.[56]
Детоубийство часто встречается у полигинных видов, таких как серые лангуры и гориллы. Взрослые самцы могут убивать зависимое потомство, которое им не принадлежит, поэтому самка вернется в течку и, таким образом, они могут произвести собственное потомство. Социальная моногамия у некоторых видов могла развиться для борьбы с этим поведением.[126] Распущенность может также снизить риск детоубийства, поскольку отцовство становится неопределенным.[127]
У приматов больше малолетний период между отлучением от груди и половым созреванием, чем у других млекопитающих аналогичного размера.[56] Некоторые приматы, такие как Galagos и новые мировые обезьяны используйте дупла деревьев для гнездования и паркуйте молодь на грядках во время кормления. Другие приматы придерживаются стратегии «верховой езды», т. Е. Несут особей на теле во время кормления. Взрослые особи могут строить или использовать места для гнездования, иногда в сопровождении молодых особей, с целью отдыха, поведение, которое вторично развилось у человекообразных обезьян.[128][129] В молодом периоде приматы более восприимчивы, чем взрослые хищничество и голодание; в это время они приобретают опыт кормления и ухода от хищников.[56] Они изучают социальные и боевые навыки, часто в процессе игры.[56] Приматы, особенно самки, живут дольше, чем другие млекопитающие того же размера.[56] это может быть частично связано с их более медленным метаболизмом.[130] В конце жизни у самок приматов, страдающих катаррином, нарушается репродуктивная функция, известная как менопауза; другие группы менее изучены.[131]
Диета и кормление


Приматы используют множество источников пищи. Было сказано, что многие характеристики современных приматов, включая людей, происходят от практики ранних предков брать большую часть своей пищи из тропического полога.[132] Большинство приматов включают в свой рацион фрукты, чтобы получить легко усваиваемые питательные вещества, в том числе углеводы и липиды для энергии.[56] Приматы в подотряде Strepsirrhini (не долгопятые просимиане) способны синтезировать Витамин С, как и большинство других млекопитающих, приматы подотряда Haplorrhini (долгопяты, обезьяны и обезьяны) утратили эту способность и нуждаются в этом витамине в своем рационе.[133]
Многие приматы обладают анатомической специализацией, которая позволяет им использовать определенную пищу, такую как фрукты, листья, жевательную резинку и т. Д. насекомые.[56] Например, листоеды, такие как обезьяны-ревуны, черно-белые колобусы и спортивные лемуры имеют расширенные пищеварительные тракты, что позволяет им поглощать питательные вещества из трудно перевариваемых листьев.[56] Мартышки, которые едят жевательную резинку, имеют сильные резец зубы, позволяющие им открывать кору деревьев, чтобы добраться до десен, и когти, а не гвозди, позволяющие им цепляться за деревья во время кормления.[56] В да-да Сочетает в себе зубы грызуна с длинным тонким средним пальцем, чтобы заполнить ту же экологическую нишу, что и дятел. Он стучит по деревьям, чтобы найти личинки насекомых, затем прогрызает отверстия в древесине и вставляет свой удлиненный средний палец, чтобы вытащить личинки.[134] У некоторых видов есть дополнительные специализации. Например, серощекий мангабей имеет толстый эмаль на зубах, что позволяет открывать твердые плоды и семена, которые другие обезьяны не могут.[56] В Gelada - единственный вид приматов, который питается в основном травой.[135]
Охота

Долгопяты единственные сохранившийся облигатный плотоядный приматы, питающиеся исключительно насекомыми, ракообразными, мелкими позвоночными и змеями (в том числе ядовитый разновидность).[136] Обезьяны капуцины может использовать различные типы растений, включая фрукты, листья, цветы, бутоны, нектар и семена, но также есть насекомых и другие беспозвоночные, птичьи яйца и мелкие позвоночные, такие как птицы, ящерицы, белки и летучие мыши.[90]
В обыкновенный шимпанзе ест всеядный плодоядный рацион питания. Он предпочитает фрукты всем остальным продуктам питания и даже ищет и ест их, когда их мало. Он также ест листья и листовые почки, семена, цветки, стебли, сердцевину, кору и смолу. Насекомые и мясо составляют небольшую часть их рациона, по оценкам, 2%.[137][138] Потребление мяса включает в себя истребление других видов приматов, таких как западный красный колобус обезьяна.[111] Бонобо - это всеядный плодоядное животное - большую часть его рациона составляют фрукты, но он дополняет их листьями, мясом из мелких позвоночные, Такие как аномалии, белки-летяги и Duikers,[139] и беспозвоночные.[140] В некоторых случаях было показано, что бонобо поедают приматов низшего порядка.[141][142]
До развития сельского хозяйства примерно 10 000 лет назад Homo sapiens использовали метод охотников-собирателей в качестве единственного средства сбора пищи. Это включало комбинирование стационарных источников пищи (таких как фрукты, зерна, клубни и грибы, личинки насекомых и водных моллюсков) с Дикая игра, на которых нужно охотиться и убивать, чтобы их можно было съесть.[143] Было высказано предположение, что люди использовали огонь для подготовки и повар еда со времен человек прямоходящий.[144] Около десяти тысяч лет назад люди развили сельское хозяйство,[145] которые существенно изменили их диету. Это изменение в диете могло также повлиять на биологию человека; с распространением молочное животноводство обеспечение нового и богатого источника пищи, ведущего к развитию способности переваривать лактоза у некоторых взрослых.[146][147]
Как добыча
Хищники приматов включают различные виды плотоядные, хищные птицы, рептилии, и другие приматы. Даже гориллы были зарегистрированы как жертвы. У хищников приматов есть различные стратегии охоты, и поэтому приматы развили несколько разных стратегий охоты. антихищные приспособления включая крипсида, тревожные звонки и моббинг. Некоторые виды имеют отдельные сигналы тревоги для различных хищников, таких как хищники, обитающие в воздухе или на земле. Хищничество могло сформировать размер группы у приматов, поскольку виды, подвергавшиеся более высокому давлению хищников, по-видимому, живут большими группами.[148]
Интеллект и познание
Приматы обладают развитыми когнитивными способностями: некоторые создают инструменты и используют их для приобретения еды и для общественных мероприятий;[149][150] некоторые могут выполнять задачи, требующие сотрудничества, влияния и ранга;[151] они осознают статус, склонны к манипуляциям и обману;[152][153] они могут распознать родственник и сородичи;[154][155] и они могут научиться использовать символы и понимать аспекты человеческого языка, включая некоторый реляционный синтаксис и концепции числа и числовой последовательности.[156][157][158] Исследования познания приматов исследуют решение проблем, память, социальное взаимодействие, теория разума, а также числовые, пространственные и абстрактные концепции.[159] Сравнительные исследования показывают тенденцию к более высокому интеллекту, переходящему от прозимианов к обезьянам Нового Света и обезьянам Старого Света, и значительно более высокие средние когнитивные способности у человекообразных обезьян.[160][161] Однако в каждой группе существует много различий (например, среди обезьян Нового Света обе паук[160] и обезьяны капуцины[161] получили высокие оценки по некоторым параметрам), а также по результатам различных исследований.[160][161]
Использование и производство инструмента


В 1960 г. Джейн Гудолл заметил шимпанзе тыкать кусочками травы в термит холмик, а затем поднял траву ко рту. После того, как он ушел, Гудолл подошла к холму и повторила то же самое, потому что не знала, что делает шимпанзе. Она обнаружила, что термиты кусали траву челюстями. Шимпанзе использовали траву как инструмент для «ловли» или «окунания» термитов.[162] Есть более ограниченные сообщения о близких родственниках бонобо, использующих инструменты в дикой природе; Утверждается, что они редко используют инструменты в дикой природе, хотя они используют инструменты так же охотно, как шимпанзе в неволе.[163] Сообщалось, что самки, как шимпанзе, так и бонобо, используют инструменты более жадно, чем самцы.[164] Орангутаны в Борнео выловить сома из небольших водоемов. Антрополог Энн Руссон видела, как несколько животных на этих лесных островах учились самостоятельно колоть сома палками, чтобы запаникованная добыча выпрыгивала из пруда и попадала в ожидающие руки орангутана.[165] Есть несколько сообщений о гориллы используя инструменты в дикой природе. Взрослая самка западная низменная горилла использовала ветку как трость, по-видимому, чтобы проверить глубину воды и помочь ей пересечь водоем. Другая взрослая самка использовала оторванный ствол небольшого куста в качестве стабилизатора во время сбора пищи, а другая использовала бревно в качестве моста.[166]
Черно-полосатый капуцин был первым приматом, не являющимся обезьяной, для которого было зарегистрировано использование инструментов в дикой природе; были замечены люди, раскалывающие орехи, кладя их на каменную наковальню и ударяя по ним другим большим камнем.[167] В Таиланде и Мьянме крабоядные макаки используйте каменные инструменты, чтобы открыть орехи, устриц и других двустворчатых моллюсков, а также различных видов морских улиток.[168] Павианы чакма используют камни как оружие; Эти павианы забивают камнями каменные стены каньона, где они спят и отступают, когда им угрожают. Камни поднимаются одной рукой и сбрасываются за борт, после чего они падают со скалы или падают прямо на дно каньона.[169]
Хотя в дикой природе лемуры не использовали инструменты, было показано, что в контролируемых условиях лемуры способны понимать функциональные свойства объектов, которых они были обучены использовать в качестве инструментов, выполняя, а также используя гаплоринги.[170]
Изготовление инструментов гораздо реже, чем использование простых инструментов, и, вероятно, представляет собой более высокое когнитивное функционирование. Вскоре после своего первого открытия использования инструментов, Гудолл наблюдала, как другие шимпанзе собирают лиственные веточки, срывают листья и используют стебли для ловли насекомых. Превращение листовой веточки в инструмент было большим открытием. До этого ученые думали, что только люди производил и использовал инструменты, и именно эта способность отделяла людей от других животных.[162] Также были замечены шимпанзе, которые делают «губки» из листьев и мха, которые всасывают воду.[171] Было замечено, что суматранские орангутаны изготавливают и используют инструменты. Они отламывают ветку дерева длиной около 30 см, отрывают ветки, перетирают один конец и затем с помощью палки выкапывают дупла деревьев для термитов.[172][173] В дикой природе мандрилы чистят уши модифицированными инструментами. Ученые засняли большой мужской мандрил на Честерский зоопарк (Великобритания) срезает ветку, очевидно, чтобы сделать ее уже, а затем, используя модифицированную палку, соскребает грязь из-под ногтей на ногах.[174] Пленные гориллы сделали множество инструментов.[175]
Выбор партнера
Выбор партнера важен для самок приматов, потому что он способствует их репродуктивному успеху. [176]. Выбор самки приматов определяется как способность самок приматов выбирать высококачественного самца для совокупления.[176] и приносит косвенные и прямые выгоды.
Прямая польза для репродуктивного успеха самки определяется способностью самца повышать шансы на выживание своего потомства, что часто обеспечивается отцовской заботой. В идеале самка предпочла бы совокупляться с самцом, который может защитить ее потомство. Хотя вклад отцов в уход за потомством со стороны мужчин, как правило, невелик и сильно варьируется в зависимости от вида приматов.[177], некоторые способы, которыми самец может заботиться о потомстве и защищать его, чтобы увеличить его шансы на выживание, включают защиту от агрессии и детоубийства со стороны других самцов.[178]. С другой стороны, косвенные выгоды - это те, которые влияют на генетическое качество потомства; следовательно, они предсказывают, что самка выберет партнера с высоким генетическим качеством.[179]. Хотя способность самок выбирать себе партнеров дает свои преимущества, важно отметить, что они не всегда могут действовать в соответствии со своими предпочтениями.
Доказательства предпочтения партнера женского пола
Одним из косвенных преимуществ выбора партнера является генетическое качество потенциального партнера-мужчины, которое может проявляться в физических характеристиках. Самки приматов отдают предпочтение определенным физическим характеристикам, и есть две гипотезы о связи между косвенными преимуществами и физическими характеристиками. Первый был разработан генетиком Рональдом Фишером, который предположил, что, когда самки выбирают самца с хорошими генами или «привлекательными чертами», они передаются сыновьям, а затем увеличивают репродуктивный успех потомства самки.[180]. Один из примеров - самки макак-резус, которые предпочитают самцов с более красными и симметричными лицами.[181][182] и, выбрав самца с красным или симметричным лицом, эти характеристики также передаются своим потомкам. Вторая гипотеза заключается в том, что связь между генетикой и физическими особенностями заключается в том, что они могут быть индикаторами физиологических качеств самца.[183][184]. В некоторых случаях украшения являются физическими индикаторами хороших генов, связанных с такими вещами, как иммунокомпетентность у мужчин.[185]. Такие гены, связанные с иммунным ответом, включают гены MHC, которые у лемуров физически проявляются в запахе, который они выпускают из своих желез, и который находится на их хвостах.[186]. В этом случае имеет смысл, что самка предпочтет определенные физические особенности, если это означает, что их потомство унаследует высококачественную иммунную систему.
Свидетельства выбора партнера женского пола
На связь между предпочтениями женщин и фактическим выбором может влиять социальная динамика группы среди женщин и мужчин. Как показано на примере верветок, самки приматов предпочитают спариваться с альфа-самцами; однако, в зависимости от социального положения самки, самец может отклонять предложения о спаривании со стороны самки.[187]. Это означает, что даже когда мужчины явно предпочитают спариваться с альфа-самцами, они не всегда могут действовать в соответствии с этим. Одна проблема, которая поднимается при этом, связана с тем, как отделить выбор самки от конкуренции между мужчиной и мужчиной, поскольку часто неясно, какой механизм создает результаты.[188]. Тем не менее, у японских макак вмешательство самца в половое взаимодействие самца и самки у японских макак не влияет на выбор самки.[189], что показывает, что в этом случае даже при соревновании самец-самец выбор самки более важен.
Экология

Нечеловеческие приматы в основном живут в тропический широты Африки, Азии и Америки. Виды, обитающие за пределами тропиков; включить Японская макака который обитает на японских островах Хонсю и Хоккайдо; то Берберийская макака который обитает в Северной Африке, и несколько видов лангуров, обитающих в Китае. Приматы обычно живут в тропические леса но также встречаются в умеренные леса, саванны, пустыни, горы и прибрежные районы.[190] Количество видов приматов в тропических районах положительно оценивается. коррелированный к количеству осадков и площади влажных лесов.[191] На долю от 25% до 40% животные, питающиеся фруктами (к масса ) в тропических лесах приматы играют важную роль экологический роль за счет распространения семян многих древесных пород.[192]
Среда обитания приматов охватывает диапазон высот: черная курносая обезьяна был найден живущим в Горы Хэндуань на высоте 4700 метров (15400 футов),[193] то горная горилла находится на высоте 4200 метров (13 200 футов), пересекая Горы Вирунга,[194] и гелада была обнаружена на высоте до 5000 м (16000 футов) в Эфиопское нагорье.[195] Некоторые виды взаимодействуют с водной средой и могут плавать или даже нырять, в том числе хоботок обезьяны, Обезьяна де Бразза и Болотная обезьяна Аллена.[196] Некоторые приматы, такие как макака резус и серые лангуры, могут эксплуатировать измененную человеком среду и даже жить в городах.[106][197]
Взаимодействие между людьми и другими приматами
Передача болезни
Тесные взаимодействия между людьми и приматами, не являющимися людьми (NHP), могут создавать пути передачи зоонозы. Вирусы, такие как Herpesviridae (особенно Вирус герпеса B ), Поксвириды, корь, Эбола, бешенство, то Марбургский вирус и вирусный гепатит может передаваться человеку; в некоторых случаях вирусы вызывают потенциально смертельные заболевания как у людей, так и у приматов.[198]
Правовой и социальный статус

Только люди признаны как человек и охраняется законом Объединенные Нации Всеобщая декларация прав человека.[b] С другой стороны, правовой статус НПЗ является предметом многочисленных споров с такими организациями, как Великая обезьяна (GAP) кампания присудить хотя бы некоторым из них законные права.[200] В июне 2008 года Испания стала первой страной в мире, признавшей права некоторых НПЗ, когда межпартийный комитет по охране окружающей среды в ее парламенте призвал страну выполнить рекомендации GAP, а именно: шимпанзе, бонобо, орангутаны, и гориллы не должны использоваться для экспериментов на животных.[201][202]
Многие виды NHP содержатся людьми в качестве домашних животных, по оценкам Allied Effort to Save Other Primates (AESOP), около 15 000 NHP живут как экзотические домашние животные в Соединенных Штатах.[203] Расширяющийся китайский средний класс в последние годы увеличил спрос на NHP как экзотических домашних животных.[204] Хотя импорт NHP для торговли домашними животными был запрещен в США в 1975 году, контрабанда все еще имеет место по США - граница Мексики с ценами от АМЕРИКАНСКИЙ ДОЛЛАР$ 3000 для обезьян до 30 000 долларов для обезьян.[205]
Приматы используются как модельные организмы в лабораториях и использовались в космические миссии.[206] Они служат служебные животные для людей с ограниченными возможностями. Обезьяны капуцины возможно обученный помогать парализованный люди; их интеллект, память и ловкость рук делают их идеальными помощниками.[207]
НПЗ хранятся в зоопарки вокруг света. Исторически зоопарки были в первую очередь формой развлечения, но в последнее время они сместили акцент на охрану, образование и исследования. GAP не настаивает на том, чтобы все NHP были выпущены из зоопарков, прежде всего потому, что рожденным в неволе приматам не хватает знаний и опыта, чтобы выжить в дикой природе в случае выпуска.[208]
Роль в научных исследованиях

Тысячи нечеловеческих приматов используются во всем мире в исследованиях из-за их психологического и физиологического сходства с людьми.[209][210] В частности, мозг и глаза NHP более близки к анатомии человека, чем у любых других животных. NHP обычно используются в доклинические испытания, нейробиология, офтальмология исследования и исследования токсичности. Макаки резус часто используются, как и другие макаки, Африканский зеленые обезьяны, шимпанзе, бабуины, беличьи обезьяны, и мартышки, как выловленные в природе, так и специально выведенные.[209][211]
В 2005 году GAP сообщил, что 1280 из 3100 NHP, живущих в неволе в Соединенных Штатах, использовались для экспериментов.[200] В 2004 г. Евросоюз в таких экспериментах использовалось около 10 000 NHP; в 2005 г. в Великобритании было проведено 4652 эксперимента на 3115 НПЗ.[212] Правительства многих стран предъявляют строгие требования к содержанию NHP в неволе. В США федеральные правила широко регулируют аспекты содержания, кормления, обогащения и разведения NHP.[213] Европейские группы, такие как Европейская коалиция за прекращение экспериментов на животных добиваются запрета на любое использование NHP в экспериментах в рамках проводимого Европейским союзом обзора законодательства об испытаниях на животных.[214]
Угрозы исчезновения

В Международный союз охраны природы (МСОП) перечисляет более трети приматов как находящиеся под угрозой исчезновения или уязвимые. Около 60% видов приматов находятся под угрозой исчезновения, в том числе: 87% видов на Мадагаскаре, 73% в Азии, 37% в Африке и 36% в Южной и Центральной Америке.[183] Кроме того, у 75% видов приматов уменьшается популяция.[183] Торговля регулируется, так как все виды перечислены СИТЕС в Приложение II., кроме 50 видов и подвидов, перечисленных в Приложение I, которые получают полную защиту от торговли.[215][216]
Общие угрозы для видов приматов включают: вырубка леса, фрагментация леса, обезьяна едет (в результате набега приматов),[217] и охота на приматов для использования в лекарствах, в качестве домашних животных и в пищу. Широкомасштабная вырубка тропических лесов широко считается процессом, который представляет наибольшую опасность для приматов.[218][219][220] Более 90% видов приматов обитают в тропических лесах.[219][221] Основная причина вырубки лесов - вырубка под сельское хозяйство, хотя коммерческие рубки пропитание лесозаготовка, добыча полезных ископаемых и строительство плотин также способствуют уничтожению тропических лесов.[221] В Индонезии большие площади равнинных лесов вырублены для увеличения пальмовое масло производства, и один анализ спутниковых снимков показал, что в 1998 и 1999 годах было потеряно 1000 Суматранские орангутаны в год в Экосистема Leuser один.[222]

Приматы с большим размером тела (более 5 кг) подвергаются повышенному риску исчезновения из-за их большей прибыльности для браконьеры по сравнению с более мелкими приматами.[221] Они позже достигают половой зрелости и имеют более длительный период между родами. Поэтому популяции восстанавливаются медленнее после того, как они были истощены браконьерством или торговлей домашними животными.[223] Данные по некоторым африканским городам показывают, что половина всего белка, потребляемого в городских районах, поступает из мясо диких животных торговля.[224] Вымирающие приматы, такие как геноны и дрель на них охотятся на уровнях, которые намного превышают устойчивые уровни.[224] Это связано с их большими размерами тела, простотой транспортировки и рентабельностью в пересчете на животное.[224] По мере того, как сельское хозяйство вторгается в лесные среды обитания, приматы питаются урожаем, нанося фермерам большие экономические потери.[225] Набеги приматов на посевы создают у местных жителей негативное впечатление о приматах, препятствуя их сохранению.[226]
Мадагаскар, где проживают пять эндемичных семейств приматов, пережил величайшее исчезновение в недавнем прошлом; Со времени человеческого поселения 1500 лет назад по крайней мере восемь классов и пятнадцать более крупных видов вымерли из-за охоты и разрушения среды обитания.[69] Среди уничтоженных приматов были Археоиндрис (лемур крупнее серебристой гориллы) и семьи Палеопропитеки и Archaeolemuridae.[69]

В Азии индуизм, буддизм и ислам запрещают есть мясо приматов; однако на приматов по-прежнему охотятся ради еды.[221] Некоторые более мелкие традиционные религии разрешают употребление мяса приматов.[227][228] Торговля домашними животными и народная медицина также увеличивают спрос на незаконную охоту.[204][229][230] В макака резус, а модельный организм, был защищен после того, как чрезмерный отлов угрожал его численности в 1960-х; программа оказалась настолько эффективной, что теперь они рассматриваются как вредители для всего их ареала.[220]
В Центральной и Южной Америке фрагментация лесов и охота - две основные проблемы для приматов. Большие участки леса сейчас редкость в Центральной Америке.[218][231] Это увеличивает количество леса, уязвимого для краевые эффекты такие как вторжение сельскохозяйственных угодий, снижение уровня влажности и изменение жизни растений.[232][233] Ограничение передвижения приводит к большему количеству инбридингов, что может вызвать пагубные последствия, приводящие к узкое место населения, в результате чего теряется значительный процент населения.[234][235]
Существует 21 примат, находящийся под угрозой исчезновения, 7 из которых остались членами МСОП "25 самых находящихся под угрозой исчезновения приматов в мире "список с 2000 года: шелковистая сифака, Лангур Делакура, то белоголовый лангур, то серые черенки, то Тонкинская курносая обезьяна, то Кросс-Ривер горилла и Суматранский орангутанг.[236] Красный колобус мисс Уолдрон был недавно объявлен вымершим, когда с 1993 по 1999 год не удалось найти никаких следов подвида.[237] С тех пор несколько охотников нашли и убили особей, но перспективы этого подвида остаются мрачными.[238]
Смотрите также
Сноски
- ^ а б Хотя монофилетические отношения между лемурами и лорисоидами широко распространено, название их клады - нет. Термин «лемуриформа» используется здесь, потому что он происходит от одной популярной таксономии, объединяющей клады зубчатых приматов в один инфраотряд и вымершие, незачищенные Adapiforms в другой, как внутри подотряд Strepsirrhini.[16][17] Однако другая популярная альтернативная таксономия помещает лорисоиды в собственном инфраотряде, Lorisiformes.[15]
- ^ Статья 6: Каждый имеет право везде быть правосубъектным.[199]
Рекомендации
- ^ а б c d е ж грамм час Гровс, К. П. (2005). Уилсон, Д.; Ридер, Д. М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Балтимор: Издательство Университета Джона Хопкинса. С. 111–184. ISBN 0-801-88221-4. OCLC 62265494.
- ^ Силкокс, Мэри Т .; Блох, Джонатан I .; Boyer, Doug M .; Честер, Стивен Г. Б.; Лопес-Торрес, Серги (2017). «Эволюционное излучение плезиадапиформ». Эволюционная антропология: проблемы, новости и обзоры. 26 (2): 74–94. Дои:10.1002 / evan.21526. ISSN 1520-6505. PMID 28429568.
- ^ «Примат». Онлайн-словарь Merriam-Webster. Мерриам-Вебстер. Получено 2008-07-21.
- ^ Книга научно-популярных. 1963. с. 257.
- ^ Анон. (1911), «Обезьяна», Британская энциклопедия, XIX (11-е изд.), Нью-Йорк: Британская энциклопедия, получено 2011-07-10
- ^ Диксон, А.Ф. (1981), Естественная история гориллы, Лондон: Weidenfeld & Nicolson, ISBN 978-0-297-77895-0
- ^ Определения парафилия отличаться; для используемого здесь см., например, Стэйс, Клайв А. (2010), «Классификация по молекулам: что это даст ботаникам-ботаникам?» (PDF), Watsonia, 28: 103–122, архивировано с оригинал (PDF) на 2011-07-26, получено 2010-02-07.
- ^ Определения монофилия отличаться; для используемого здесь см., например, Мишлер, Брент Д. (2009), «Виды не являются уникально реальными биологическими объектами», в Ayala, F.J. & Arp, R. (eds.), Современные дискуссии по философии биологии, стр. 110–122, Дои:10.1002 / 9781444314922.ch6, ISBN 978-1-4443-1492-2.
- ^ а б Картмилл, М .; Смит, Ф. Х. (2011). Человеческое происхождение. Джон Вили и сыновья. ISBN 978-1-118-21145-8.
- ^ а б Гровс, С. П. (2001). Таксономия приматов. Пресса Смитсоновского института. ISBN 1-56098-872-X.
- ^ Бентон 2005, п. 371.
- ^ Бентон 2005 С. 378–380.
- ^ а б Миттермайер, Р.; Ganzhorn, J .; Konstant, W .; Glander, K .; Tattersall, I .; Groves, C .; Rylands, A .; Hapke, A .; Ratsimbazafy, J .; Мэр, М .; Louis, E .; Rumpler, Y .; Schwitzer, C .; Rasoloarison, R. (декабрь 2008 г.). «Разнообразие лемуров на Мадагаскаре» (PDF). Международный журнал приматологии. 29 (6): 1607–1656. Дои:10.1007 / s10764-008-9317-у. S2CID 17614597.
- ^ а б Райлендс, А. Б. и Миттермайер, Р. А. (2009). «Разнообразие приматов Нового Света (Platyrrhini)». В Гарбер, П. А .; Эстрада, А .; Bicca-Marques, J.C .; Heymann, E. W. & Стриер, К. (ред.). Южноамериканские приматы: сравнительные перспективы в изучении поведения, экологии и охраны природы. Springer. ISBN 978-0-387-78704-6.
- ^ а б Хартвиг 2011 С. 20–21.
- ^ Салай и Делсон 1980, п. 149.
- ^ Картмилл 2010, п. 15.
- ^ Линней, К. (1758). Sistema naturae per regna tria Naturae, классы secundum, обыкновенные, роды, виды, cum characteribus diffis, synonimis locis. Томус I. Impensis прямой. Лаурентий Сальвии, Холмия. С. 20–32.
- ^ Линней, К. (1735). Sistema naturae sive regna tria Naturae systematice proposita по классам, порядкам, родам и видам. apud Theodorum Haak, Lugduni Batavorum. стр. s.p.
- ^ Блейнвилл, Х. (1839). "Новая классификация маммфер". Французские Анналы и Этранжеры анатомии и физиологии Appliquées à la Médicine et à l'Histoire Naturelle, 3. С. 268–269.
- ^ Торингтон, Р. В. и Андерсон, С. (1984). «Приматы». В Андерсон, С. и Джонс, Дж. К. (ред.). Отряды и семейства современных млекопитающих мира. Нью-Йорк: Джон Уайли и сыновья. стр.187–217. ISBN 978-0-471-08493-8.
- ^ Маккенна, М.С. и Белл, С.К. (1997). Классификация млекопитающих: выше уровня вида. Нью-Йорк: издательство Колумбийского университета. п. 631. ISBN 0-231-11013-8.
- ^ Стриер, К. (2007). Поведенческая экология приматов (Третье изд.). С. 50–53. ISBN 978-0-205-44432-8.
- ^ Janečka, J. E .; Miller, W .; Pringle, T. H .; Wiens, F .; Zitzmann, A .; Helgen, K. M .; Springer, M. S .; Мерфи, У. Дж. (2 ноября 2007 г.). «Молекулярные и геномные данные идентифицируют ближайших живых родственников приматов». Наука. 318 (5851): 792–794. Bibcode:2007Sci ... 318..792J. Дои:10.1126 / science.1147555. PMID 17975064. S2CID 12251814.
- ^ Кавана, М. (1983). Полное руководство по обезьянам, обезьянам и другим приматам. Нью-Йорк: Viking Press. стр.18. ISBN 0-670-43543-0.
- ^ Маккенна, М.С. и Белл, С.К. (1997). Классификация млекопитающих выше видового уровня. Нью-Йорк: издательство Колумбийского университета. п. 329. ISBN 0-231-11012-X.
- ^ Williams, B.A .; Kay, R.F .; Кирк, E.C. (2010). «Новые взгляды на происхождение антропоидов». Труды Национальной академии наук Соединенных Штатов Америки. 107 (11): 4797–4804. Bibcode:2010PNAS..107.4797W. Дои:10.1073 / pnas.0908320107. ЧВК 2841917. PMID 20212104.
- ^ Станьон, Роско; Springer, Mark S .; Мередит, Роберт В .; Гейтси, Джон; Эмерлинг, Кристофер А .; Парк, Джонг; Rabosky, Daniel L .; Стадлер, Таня; Штайнер, Синтия; Райдер, Оливер А .; Janečka, Jan E .; Фишер, Коллин А.; Мерфи, Уильям Дж. (2012). "Макроэволюционная динамика и историческая биогеография диверсификации приматов на основе суперматрицы видов". PLOS ONE. 7 (11): e49521. Bibcode:2012PLoSO ... 749521S. Дои:10.1371 / journal.pone.0049521. ISSN 1932-6203. ЧВК 3500307. PMID 23166696.
- ^ Джеймсон, Натали М .; Хоу, Чжо-Чэн; Стернер, Кирстин Н .; Weckle, Эми; Гудман, Моррис; Steiper, Michael E .; Уайлдман, Дерек Э. (сентябрь 2011 г.). «Геномные данные опровергают гипотезу о кладе просимиевых приматов». Журнал эволюции человека. 61 (3): 295–305. Дои:10.1016 / j.jhevol.2011.04.004. ISSN 0047-2484. PMID 21620437.
- ^ Поцци, Лука; Ходжсон, Джейсон А .; Баррелл, Эндрю С .; Стернер, Кирстин Н .; Raaum, Ryan L .; Дисотелл, Тодд Р. (июнь 2014 г.). «Филогенетические отношения приматов и даты дивергенции, выведенные из полных митохондриальных геномов». Молекулярная филогенетика и эволюция. 75: 165–183. Дои:10.1016 / j.ympev.2014.02.023. ISSN 1055-7903. ЧВК 4059600. PMID 24583291.
- ^ Станьон, Роско; Финстермайер, Кнут; Зиннер, Дитмар; Брамейер, Маркус; Мейер, Матиас; Кройц, Ева; Хофрайтер, Майкл; Роос, Кристиан (16 июля 2013 г.). «Митогеномная филогения живых приматов». PLOS ONE. 8 (7): e69504. Bibcode:2013PLoSO ... 869504F. Дои:10.1371 / journal.pone.0069504. ISSN 1932-6203. ЧВК 3713065. PMID 23874967.
- ^ Уильямс, Б. А .; Kay, R.F .; Кирк, Э. К. (2010). «Новые взгляды на происхождение антропоидов». Труды Национальной академии наук Соединенных Штатов Америки. 107 (11): 4797–4804. Bibcode:2010PNAS..107.4797W. Дои:10.1073 / pnas.0908320107. ЧВК 2841917. PMID 20212104.
- ^ Miller, E. R .; Gunnell, G.F .; Мартин, Р. Д. (2005). "Глубокое время и поиски истоков антропоидов" (PDF). Американский журнал физической антропологии. 128: 60–95. Дои:10.1002 / ajpa.20352. PMID 16369958.
- ^ а б Чаттерджи, Хелен Дж; Хо, Саймон Ю.В.; Барнс, Ян; Гровс, Колин (27 октября 2009 г.). «Оценка филогении и времени дивергенции приматов с использованием суперматричного подхода». BMC Эволюционная биология. 9 (1): 259. Дои:10.1186/1471-2148-9-259. ЧВК 2774700. PMID 19860891.
- ^ О'Лири, М. А .; и другие. (8 февраля 2013 г.). «Предок плацентарных млекопитающих и пост-K-Pg-излучение плаценты». Наука. 339 (6120): 662–667. Bibcode:2013Наука ... 339..662O. Дои:10.1126 / science.1229237. PMID 23393258. S2CID 206544776.
- ^ Ли, М. (сентябрь 1999 г.). "Молекулярные часы калибровки и даты расхождения многоклеточных животных". Журнал молекулярной эволюции. 49 (3): 385–391. Bibcode:1999JMolE..49..385L. Дои:10.1007 / PL00006562. PMID 10473780. S2CID 1629316.
- ^ «Ученые отодвигают происхождение приматов с 65 до 85 миллионов лет назад». Science Daily. Получено 2008-10-24.
- ^ Tavaré, S .; Marshall, C.R .; Will, O .; Soligo, C .; Мартин Р.Д. (18 апреля 2002 г.). «Использование летописи окаменелостей для оценки возраста последнего общего предка современных приматов». Природа. 416 (6882): 726–729. Bibcode:2002Натура 416..726Т. Дои:10.1038 / 416726a. PMID 11961552. S2CID 4368374.
- ^ Klonisch, T .; Froehlich, C .; Тетенс, Ф .; Фишер, Б .; Хомбах-Клониш, С. (2001). «Молекулярное ремоделирование членов семейства релаксинов во время эволюции приматов». Молекулярная биология и эволюция. 18 (3): 393–403. Дои:10.1093 / oxfordjournals.molbev.a003815. PMID 11230540. Получено 2008-08-22.
- ^ а б c d е ж Horvath, J .; и другие. (2008). «Разработка и применение филогеномного инструментария: решение эволюционной истории мадагаскарских лемуров». Геномные исследования. 18 (3): 489–499. Дои:10.1101 / гр.7265208. ЧВК 2259113. PMID 18245770.
- ^ а б c d Продавцы, Билл (2000-10-20). "Эволюция примаса" (PDF). Эдинбургский университет. С. 13–17. Архивировано из оригинал (PDF) на 2008-10-29. Получено 2008-10-23.
- ^ а б c d Хартвиг, В. (2007). «Эволюция примаса». В Campbell, C .; Fuentes, A .; MacKinnon, K .; Panger, M .; Бердер, С. (ред.). Приматы в перспективе. Издательство Оксфордского университета. С. 13–17. ISBN 978-0-19-517133-4.
- ^ Уильямс, Б. А .; Kay, R.F .; Кристофер Кирк, E .; Росс, К. Ф. (2010). "Darwinius masillae Strepsirrhine - ответ Франзену и другие. (2009)" (PDF). Журнал эволюции человека. 59 (5): 567–573, обсуждение 573–9. Дои:10.1016 / j.jhevol.2010.01.003. PMID 20188396. Архивировано из оригинал (PDF) на 2013-05-17.
- ^ Чочон Р. и Флигл Дж. (1987). Эволюция приматов и происхождение человека. Менло-Парк, Калифорния: Бенджамин / Каммингс. п. 72. ISBN 978-0-202-01175-2.
- ^ а б Гарбут, Н. (2007). Млекопитающие Мадагаскара, полное руководство. A&C Black Publishers. С. 85–86. ISBN 978-0-300-12550-4.
- ^ а б Миттермайер, Р.А.; и другие. (2006). Лемуры Мадагаскара (2-е изд.). Консервейшн Интернэшнл. С. 23–26. ISBN 1-881173-88-7.
- ^ Шекелле, М. (2005). Эволюционная биология долгопятов. Архивировано из оригинал на 2008-09-07. Получено 2008-08-22.
- ^ Schmidt, T .; и другие. (3 мая 2005 г.). «Быстрая электростатическая эволюция в сайте связывания цитохрома с на цитохром с оксидазе у антропоидных приматов». Труды Национальной академии наук Соединенных Штатов Америки. 102 (18): 6379–6384. Bibcode:2005ПНАС..102.6379С. Дои:10.1073 / pnas.0409714102. ЧВК 1088365. PMID 15851671.
- ^ Уэйд, Лиззи (5 июня 2013 г.). «Ранний примат весил меньше унции». ScienceNow. Архивировано из оригинал на 2013-06-08. Получено 2013-06-07.
- ^ Кей, Р. Ф. (2012). «Доказательства азиатского происхождения стеблевых антропоидов». Труды Национальной академии наук Соединенных Штатов Америки. 109 (26): 10132–10133. Bibcode:2012ПНАС..10910132К. Дои:10.1073 / pnas.1207933109. ЧВК 3387095. PMID 22699505.
- ^ Chaimanee, Y .; Chavasseau, O .; Beard, K. C .; Kyaw, A. A .; Soe, A. N .; Sein, C .; Lazzari, V .; Marivaux, L .; Марандат, Б .; Swe, M .; Rugbumrung, M .; Lwin, T .; Валентин, X .; Зин-Маунг-Маунг-Тейн; Jaeger, J. -J. (2012). «Примат позднего среднего эоцена из Мьянмы и начальная антропоидная колонизация Африки». Труды Национальной академии наук Соединенных Штатов Америки. 109 (26): 10293–10297. Bibcode:2012PNAS..10910293C. Дои:10.1073 / pnas.1200644109. ЧВК 3387043. PMID 22665790.
- ^ Marivaux, L .; и другие. (2005-06-14). «Антропоидные приматы из олигоцена Пакистана (Bugti Hills): данные о ранней антропоидной эволюции и биогеографии». Труды Национальной академии наук Соединенных Штатов Америки. 102 (24): 8436–8441. Bibcode:2005ПНАС..102.8436М. Дои:10.1073 / pnas.0503469102. ЧВК 1150860. PMID 15937103.
- ^ Шраго, К. И Руссо, C.A.M. (2003). «Время появления обезьян Нового Света» (Перепечатка PDF). Молекулярная биология и эволюция. 20 (10): 1620–1625. Дои:10.1093 / molbev / msg172. PMID 12832653.
- ^ Хоул, А. (1999). «Происхождение платиррий: оценка сценария Антарктики и модели плавучего острова». Американский журнал физической антропологии. 109 (4): 541–559. Дои:10.1002 / (SICI) 1096-8644 (199908) 109: 4 <541 :: AID-AJPA9> 3.0.CO; 2-N. PMID 10423268.
- ^ Эндрюс, П. и Келли, Дж. (2007). "Расселения обезьян в среднем миоцене". Folia Primatologica. 78 (5–6): 328–343. Дои:10.1159/000105148. PMID 17855786. S2CID 19293586.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты Стриер, К. (2007). Поведенческая экология приматов (3-е изд.). Аллин и Бэкон. С. 7, 64, 71, 77, 182–185, 273–280, 284, 287–298. ISBN 978-0-205-44432-8.
- ^ Pough, F.W .; Janis, C.M .; Хайзер, Дж. Б. (2005) [1979]. «Эволюция приматов и появление людей». Позвоночные животные (7-е изд.). Пирсон. п. 650. ISBN 0-13-127836-3.
- ^ Теназа Р. (1984). «Песни гиббонов» (Hylobates lar × H. muelleri)". Американский журнал приматологии. 8 (3): 249–253. Дои:10.1002 / ajp.1350080307. PMID 31986810. S2CID 84957700.
- ^ а б Бернстейл, И. С. (1966). «Встречающийся в природе гибрид приматов». Наука. 154 (3756): 1559–1560. Bibcode:1966Sci ... 154.1559B. Дои:10.1126 / science.154.3756.1559. PMID 4958933. S2CID 85898043.
- ^ Сугавара, К. (январь 1979 г.). "Социологическое исследование дикой группы гибридных бабуинов между Папио анубис и P. hamadryas в долине Аваш, Эфиопия ". Приматы. 20 (1): 21–56. Дои:10.1007 / BF02373827. S2CID 23061688.
- ^ Jolly, C.J .; Вулли-Баркер, Тэмсин; и другие. (1997). «Межродовые гибридные бабуины». Международный журнал приматологии. 18 (4): 597–627. Дои:10.1023 / А: 1026367307470. S2CID 27900830.
- ^ Лю, Чжэнь; и другие. (24 января 2018 г.). «Клонирование макак с помощью переноса ядер соматических клеток». Клетка. 172 (4): 881–887.e7. Дои:10.1016 / j.cell.2018.01.020. PMID 29395327. Получено 24 января 2018.
- ^ Нормил, Деннис (24 января 2018 г.). «Эти близнецы-обезьяны - первые клоны приматов, созданные методом, разработанным Долли». Наука. Дои:10.1126 / science.aat1066. Получено 24 января 2018.
- ^ Сираноски, Дэвид (24 января 2018 г.). «Первые обезьяны, клонированные с помощью техники, которая сделала овечку Долли - китайские ученые создали клонированных приматов, которые могут произвести революцию в исследованиях болезней человека». Природа. 553 (7689): 387–388. Bibcode:2018Натура.553..387C. Дои:10.1038 / d41586-018-01027-z. PMID 29368720.
- ^ Бриггс, Хелен (24 января 2018 г.). «Первые клоны обезьяны созданы в китайской лаборатории». Новости BBC. Получено 24 января 2018.
- ^ «Ученые успешно клонируют обезьян. На очереди - люди?». Нью-Йорк Таймс. Ассошиэйтед Пресс. 24 января 2018 г.. Получено 24 января 2018.
- ^ а б c d Pough, F.W .; Janis, C.M .; Хайзер, Дж. Б. (2005) [1979].«Характеристики приматов». Позвоночные животные (7-е изд.). Пирсон. п. 630. ISBN 0-13-127836-3.
- ^ а б Айелло, Л. и Дин, К. (1990). Введение в эволюционную анатомию человека. Академическая пресса. п. 193. ISBN 0-12-045590-0.
- ^ а б c d е ж «Примат». Энциклопедия Britannica Online. Encyclopdia Britannica, Inc. 2008. Получено 2008-07-21.
- ^ а б c d е ж Майерс, П. (1999). ""Приматы "(Он-лайн)". Сеть разнообразия животных. Получено 2008-06-03.
- ^ Кэмпбелл Б. Г. и Лой Дж. Д. (2000). Появление человечества (8-е изд.). Аллин и Бэкон. п. 85. ISBN 0-673-52364-0.
- ^ а б Уайт, Т. и Казлев, А. (2008-01-08). «Архонта: Приматы». Палеос. Архивировано из оригинал на 2008-05-12. Получено 2008-06-03.
- ^ а б c d е ж Макдональд, Дэвид (2006). «Приматы». Энциклопедия млекопитающих. The Brown Reference Group plc. С. 282–307. ISBN 0-681-45659-0.
- ^ Ash, M. M .; Nelson, S.J .; Уилер, Р. К. (2003). Стоматологическая анатомия, физиология и окклюзия Уиллера. W.B. Сондерс. п.12. ISBN 978-0-7216-9382-8.
- ^ Гарбер PA, Rehg JA (ноябрь 1999 г.). «Экологическая роль цепкого хвоста у белолицых капуцинов (Cebus capucinus)». Американский журнал физической антропологии. 110 (3): 325–39. Дои:10.1002 / (SICI) 1096-8644 (199911) 110: 3 <325 :: AID-AJPA5> 3.0.CO; 2-D. PMID 10516564.
- ^ Руссо Г.А., Янг Дж. В. (ноябрь 2011 г.). «Рост хвоста отслеживает онтогенез использования цепкого хвоста у обезьян капуцинов (Cebus albifrons и C. apella)». Американский журнал физической антропологии. 146 (3): 465–73. Дои:10.1002 / ajpa.21617. PMID 21953012.
- ^ Фридерун Анкель-Симонс (27 июля 2010 г.). Анатомия приматов: введение. Академическая пресса. ISBN 978-0-08-046911-9.
- ^ а б c Линденфорс, Патрик; Туллберг, Биргитта С. (1998). «Филогенетический анализ эволюции размеров приматов: последствия полового отбора». Биологический журнал Линнеевского общества. 64 (4): 413–447. Дои:10.1111 / j.1095-8312.1998.tb00342.x. ISSN 0024-4066.
- ^ Раллс, К. (1976). «Млекопитающие, у которых самки крупнее самцов». Ежеквартальный обзор биологии. 51 (2): 245–76. Дои:10.1086/409310. PMID 785524. S2CID 25927323.
- ^ Линдстедтанд и Бойс; Бойс, Марк С. (июль 1985 г.). «Сезонность, выносливость при голодании и размер тела млекопитающих». Американский натуралист. 125 (6): 873. Дои:10.1086/284385. S2CID 84308684.
- ^ Торен, Сандра; Линденфорс, Патрик; Каппелер, Питер М. (2006). «Филогенетический анализ диморфизма у приматов: доказательства более сильного отбора по размеру собак, чем по размеру тела». Американский журнал физической антропологии. 130 (1): 50–59. Дои:10.1002 / ajpa.20321. ISSN 0002-9483. PMID 16345072.
- ^ Фриш, Дж. Э. (1963). «Половые различия клыков гиббона (Hylobates lar)". Приматы. 4 (2): 1–10. Дои:10.1007 / BF01659148. S2CID 189798134.
- ^ Кей, Р.Ф. (1975). «Функциональные адаптации коренных зубов приматов». Американский журнал физической антропологии. 43 (2): 195–215. Дои:10.1002 / ajpa.1330430207. PMID 810034.
- ^ Крук, Дж. Х. (1972). «Половой отбор, диморфизм и социальная организация у приматов». В Кэмпбелле, Б. Г. (ред.). Половой отбор и происхождение человека. Алдин Транзакция. стр.246. ISBN 978-0-202-02005-1.
- ^ а б Cheverud, J.M .; Dow, M. M .; Leutenegger, W. (ноябрь 1985 г.). «Количественная оценка филогенетических ограничений в сравнительном анализе: половой диморфизм массы тела среди приматов». Эволюция. 39 (6): 1335–1351. Дои:10.2307/2408790. JSTOR 2408790. PMID 28564267.
- ^ Leutenegger, W .; Чеверуд, Дж. М. (1982). «Корреляты полового диморфизма у приматов: экологические и размерные переменные». Международный журнал приматологии. 3 (4): 387–402. Дои:10.1007 / BF02693740. S2CID 38220186.
- ^ а б Плавкан, Дж. М. (2001). «Половой диморфизм в эволюции приматов». Американский журнал физической антропологии. 33: 25–53. Дои:10.1002 / ajpa.10011. PMID 11786990.
- ^ О'Хиггинс, П .; Коллард, М. (2002). «Половой диморфизм и рост лица у папиониновых обезьян». Журнал зоологии. 257 (2): 255–72. Дои:10.1017 / S0952836902000857.
- ^ Суссман, Р. В. (1999). Экология приматов и социальная структура Том 1: Лори, лемуры и долгопяты. Нидхэм-Хайтс, Массачусетс: Издательство Pearson Custom Publishing & Prentice Hall. стр.78, 89–90, 108, 121–123, 233. ISBN 0-536-02256-9.
- ^ а б c d е Суссман, Р. В. (2003). Экология приматов и социальная структура, Том 2: Обезьяны Нового Света (Пересмотренное первое издание). Нидхэм-Хайтс, Массачусетс: Издательство Pearson Custom Publishing & Prentice Hall. С. 77–80, 132–133, 141–143. ISBN 0-536-74364-9.
- ^ Glazier, S.D .; Цветочный день, К. А. (2003). Избранные чтения по антропологии религии: теоретико-методологические очерки. Издательская группа "Гринвуд". п. 53. ISBN 9780313300905.
- ^ Arrese, C.A .; Oddy, Элисон Y .; и другие. (2005). "Топография конуса и спектральная чувствительность у двух потенциально трехцветных сумчатых, квокка (Сетоникс брахюрус) и quenda (Isoodon obesulus)". Труды Королевского общества B. 272 (1565): 791–6. Дои:10.1098 / rspb.2004.3009. ЧВК 1599861. PMID 15888411.
- ^ Bowmaker, J. K .; Astell, S .; Хант, Д. М .; Моллон, Дж. Д. (1991). «Фоточувствительные и фотостабильные пигменты сетчатки глаза обезьян Старого Света» (PDF). Журнал экспериментальной биологии. 156 (1): 1–19. ISSN 0022-0949. PMID 2051127. Получено 2008-06-16.
- ^ а б Сурридж, А. К. и Д. Осорио (2003). «Эволюция и селекция трехцветного зрения у приматов». Тенденции в экологии и эволюции. 18 (4): 198–205. Дои:10.1016 / S0169-5347 (03) 00012-0.
- ^ Лукас, П. У .; Dominy, N.J .; Riba-Hernandez, P .; Стоунер, К. Э .; Yamashita, N .; Loría-Calderón, E .; Petersen-Pereira, W .; Rojas-Durán, Y .; Salas-Pena, R .; Солис-Мадригал, С .; Osorio, D .; Дарвелл, Б. У. (2003). «Эволюция и функция рутинного трихроматического зрения у приматов». Эволюция. 57 (11): 2636–43. Дои:10.1554/03-168. PMID 14686538. S2CID 739130.
- ^ Goldberg, T. L .; Wrangham, R. W. (сентябрь 1997 г.). «Генетические корреляты социального поведения у диких шимпанзе: данные митохондриальной ДНК». Поведение животных. 54 (3): 559–70. Дои:10.1006 / anbe.1996.0450. PMID 9299041. S2CID 18223362.
- ^ Фиоре, А. Д. и Кэмпбелл, К. Дж. (2007). «Ателины». В Campbell, C.J .; Fuentes, A .; MacKinnon, K. C .; Пангер М. и Бердер С. К. (ред.). Приматы в перспективе. Издательство Оксфордского университета. п. 175. ISBN 978-0-19-517133-4.
- ^ Бартлетт, Т.К. (2007). «Гилобатиды». В Campbell, C.J .; Fuentes, A .; MacKinnon, K. C .; Panger, M .; Бердер, С. К. (ред.). Приматы в перспективе. Издательство Оксфордского университета. п. 283. ISBN 978-0-19-517133-4.
- ^ Винс, Франк (2002). Поведение и экология диких медлительных лори (Nycticebus coucang): социальная организация, система ухода за младенцами и диета (PDF) (Кандидатская диссертация). Байройтский университет. С. 31–32. Архивировано из оригинал (PDF) 9 марта 2012 г.
- ^ Нотт, К. Д. и Каленберг, С. М. (2007). «Орангутаны в перспективе». В Campbell, C.J .; Fuentes, A .; MacKinnon, K. C .; Пангер М. и Бердер С. К. (ред.). Приматы в перспективе. Издательство Оксфордского университета. п. 294. ISBN 978-0-19-517133-4.
- ^ Уоттс Д. П. (1996). «Сравнительная социо-экология горилл». В McGrew W. C .; Марчант Л. Ф .; Нисида, Т. (ред.). Общества великих обезьян. Кембридж (Англия: Cambridge Univ Press. Стр. 16–28. ISBN 978-0521555364.
- ^ а б Шарпантье MJ, Widdig A, Alberts SC (декабрь 2007 г.). «Инбридинговая депрессия у нечеловеческих приматов: исторический обзор использованных методов и эмпирических данных». Американский журнал приматологии. 69 (12): 1370–86. Дои:10.1002 / ajp.20445. PMID 17486606. S2CID 46626761.
- ^ Раллс К., Баллоу Дж. (1982). «Влияние инбридинга на младенческую смертность приматов в неволе» (PDF). Международный журнал приматологии. 3 (4): 491–505. Дои:10.1007 / BF02693747. S2CID 10954608.
- ^ Констебль, Дж. Л .; Эшли, М. В .; Goodall, J .; Пусей, А. Э. (май 2001 г.). «Неинвазивное установление отцовства у шимпанзе Гомбе». Молекулярная экология. 10 (5): 1279–300. Дои:10.1046 / j.1365-294X.2001.01262.x. PMID 11380884. S2CID 46604532.
- ^ а б c d е Роу, Н. (1996). Иллюстрированный путеводитель по живым приматам. Pogonias Press. стр.4, 139, 143, 15 185, 223. ISBN 0-9648825-0-7.
- ^ Кузин, Иэн Д .; Лайдр, Марк Э. (август 2009 г.). «Деление-термоядерные популяции». Текущая биология. 19 (15): R633 – R635. Дои:10.1016 / j.cub.2009.05.034. ISSN 0960-9822. PMID 19674541. S2CID 13549970.
- ^ а б Pough, F.W .; Janis, C.M .; Хайзер, Дж. Б. (2005) [1979]. «Общества приматов». Позвоночные животные (7-е изд.). Пирсон. С. 621–623. ISBN 0-13-127836-3.
- ^ Смэтс, Б., Чейни, Д. Сейфарт, Р.М., Рэнгэм, Р.В., и Струхсакер, Т.Т. (редакторы) (1987). Общества приматов. Чикаго: Издательство Чикагского университета, где можно найти статьи о структуре и функциях различных обществ приматов.
- ^ Шульц, С. & Томсетт, С. (2007). «Взаимодействие между африканскими коронованными орлами и их жертвенным сообществом». В McGraw, W .; Zuberbuhler, K. & Noe, R. (ред.). Обезьяны леса Тай, африканское сообщество приматов. Издательство Кембриджского университета. п. 181. ISBN 978-0-521-81633-5.
- ^ а б Бшары, Р. (2007). «Взаимодействие между красными обезьянами колобус и шимпанзе». В McGraw, W .; Zuberbuhler, K .; Ноэ, Р. (ред.). Обезьяны леса Тай, африканское сообщество приматов. Издательство Кембриджского университета. С. 155–170. ISBN 978-0-521-81633-5.
- ^ Стэнфорд, К. (1998). Шимпанзе и красный колобус: экология хищника и добычи. Издательство Гарвардского университета. С. 130–138, 233. ISBN 0-674-00722-0.
- ^ а б Боински, С. (2000). «Социальные манипуляции внутри и между войсками опосредуют движение группы приматов». In Boinski, S .; Гарбер, П. (ред.). В движении: как и почему животные путешествуют группами. Издательство Чикагского университета. С. 447–448. ISBN 0-226-06340-2.
- ^ Liman, E. R .; Иннан, Х. (2003). «Ослабленное избирательное давление на важный компонент трансдукции феромона в эволюции приматов» (PDF). Труды Национальной академии наук Соединенных Штатов Америки. 100 (6): 3328–3332. Bibcode:2003ПНАС..100.3328Л. Дои:10.1073 / pnas.0636123100. ЧВК 152292. PMID 12631698. Получено 2008-07-23.
- ^ Egnor, R .; Miller, C .; Хаузер, доктор медицины (2004). "Нечеловеческое общение приматов" (PDF). Энциклопедия языка и лингвистики (2-е изд.). Эльзевир. ISBN 0-08-044299-4. Архивировано из оригинал (PDF) на 2008-09-10.
- ^ а б Pollick, A. S .; де Ваал, Ф. Б. М. (2007). «Жесты обезьяны и эволюция языка». Труды Национальной академии наук. 104 (19): 8184–8189. Bibcode:2007ПНАС..104.8184П. Дои:10.1073 / pnas.0702624104. ЧВК 1876592. PMID 17470779.
- ^ Берроуз, А. М. (2008). «Мускулатура мимики у приматов и ее эволюционное значение». BioEssays. 30 (3): 212–225. Дои:10.1002 / bies.20719. PMID 18293360. S2CID 205478149.
- ^ Ramsier, M.A .; Cunningham, A.J .; Moritz, G.L .; Finneran, J.J .; Williams, C.V .; Ong, P.S .; Gursky-Doyen, S.L .; Домини, Нью-Джерси (2012). «Общение приматов в чистом ультразвуке». Письма о биологии. 8 (4): 508–511. Дои:10.1098 / рсбл.2011.1149. ЧВК 3391437. PMID 22319094.
- ^ "Черная обезьяна-ревуна". Смитсоновский национальный зоопарк и институт биологии сохранения. Получено 2016-07-10.
- ^ Kelemen, G .; Саде, Дж. (1960). «Голосовой орган Воющей обезьяны (Alouatta palliata)". Журнал морфологии. 107 (2): 123–140. Дои:10.1002 / jmor.1051070202. PMID 13752246. S2CID 30539292.
- ^ да Кунья, Р. Г. Т .; Бирн, Р. (2006). «Рев черных обезьян-ревунов (Alouatta caraya): свидетельство функции в межгрупповом интервале». Поведение. 143 (10): 1169–1199. Дои:10.1163/156853906778691568. JSTOR 4536401.
- ^ Seyfarth, R.M .; Cheney, D. L .; Марлер, Питер (1980). «Тревожные звонки обезьян Vervet: семантическое общение в свободном примате». Поведение животных. 28 (4): 1070–1094. Дои:10.1016 / S0003-3472 (80) 80097-2. S2CID 53165940.
- ^ Fitch, W. T .; де Бур, В .; Mathur, N .; Газанфар, А. А. (2016). "Голосовые тракты обезьяны готовы к речи". Достижения науки. 2 (12): e1600723. Bibcode:2016SciA .... 2E0723F. Дои:10.1126 / sciadv.1600723. ЧВК 5148209. PMID 27957536.
- ^ Boë L.-J .; Berthommier, F .; Legou, T .; Captier, G .; Kemp, C .; Sawallis, T. R .; и другие. (2017). "Свидетельства голосовой протосистемы у бабуина (Папио папио) Предлагает предшественников речи до гоминина ». PLOS ONE. 12 (1): e0169321. Bibcode:2017PLoSO..1269321B. Дои:10.1371 / journal.pone.0169321. ЧВК 5226677. PMID 28076426.
- ^ Аркади, AC. (Август 2000 г.). «Голосовая реакция у самцов диких шимпанзе: значение для эволюции языка». J Hum Evol. 39 (2): 205–23. Дои:10.1006 / jhev.2000.0415. PMID 10968929. S2CID 7403772.
- ^ Опи, Кристофер; Аткинсон, Квентин Д .; Dunbarc, Robin I.M .; Шульц, Сюзанна (2013). «Детоубийство мужского пола приводит к социальной моногамии у приматов». Труды Национальной академии наук Соединенных Штатов Америки. 110 (33): 13328–13332. Bibcode:2013PNAS..11013328O. Дои:10.1073 / pnas.1307903110. ЧВК 3746880. PMID 23898180.
- ^ De Ruiter, Jan R .; Ван Хофф, Ян А. Р. А. М. И Шеффран, Вольфганг (1994). «Социальные и генетические аспекты отцовства у диких длиннохвостых макак (Macaca fascicularis)". Поведение. 129 (3–4): 203–24. Дои:10.1163 / 156853994x00613. JSTOR 4535195.
- ^ Каппелер, Питер М. (1998). «Гнезда, ямы в деревьях и эволюция историй жизни приматов». Американский журнал приматологии. 46 (1): 7–33. Дои:10.1002 / (SICI) 1098-2345 (1998) 46: 1 <7 :: AID-AJP3> 3.0.CO; 2- #. PMID 9730211.
- ^ Росс, Кэролайн (1991). «Парк или поездка? Эволюция вынашивания младенцев у приматов». Международный журнал приматологии. Kluwer Academic Publishing. 22 (5): 749–771. Дои:10.1023 / А: 1012065332758. S2CID 25301078.
- ^ Минц, Зоя (14 января 2014 г.). «Люди и приматы сжигают на 50 процентов меньше калорий каждый день, чем другие млекопитающие». www.ibtimes.com. IBT Media Inc. Получено 2014-01-14.
- ^ Уокер М.Л., Херндон Дж. Г.; Херндон (2008). "Менопауза у нечеловеческих приматов?". Биология размножения. 79 (3): 398–406. Дои:10.1095 / биолрепрод.108.068536. ЧВК 2553520. PMID 18495681.
- ^ Милтон, К. (1993). "Диета и эволюция приматов" (PDF). Scientific American. Vol. 269 нет. 2. С. 86–93. Bibcode:1993SciAm.269b..86M. Дои:10.1038 / scientificamerican0893-86. PMID 8351513.
- ^ Pollock, J. I .; Муллин, Р. Дж. (1986). "Биосинтез витамина С в просимианах: доказательства антропоидного сродства Tarsius". Американский журнал физической антропологии. 73 (1): 65–70. Дои:10.1002 / ajpa.1330730106. PMID 3113259. Архивировано из оригинал на 2012-06-28.
- ^ Milliken, G.W .; Ward, J. P .; Эриксон, К. Дж. (1991). "Независимый контроль пальцев при кормлении ай-ай (Daubentonia madagascariensis)". Folia Primatologica. 56 (4): 219–224. Дои:10.1159/000156551. PMID 1937286.
- ^ Хиллер, К. (2000). "Theropithecus gelada". Сеть разнообразия животных. Получено 2008-08-08.
- ^ Wright, P .; Simmons, E .; Гурский, С. (2003). "Вступление". In Wright, P .; Simmons, E .; Гурски, С. (ред.). Долгопяты прошлого, настоящего и будущего. Издательство Университета Рутгерса. п. 1. ISBN 0-8135-3236-1.
- ^ Гудолл, Джейн (1986). Шимпанзе Гомбе: модели поведения. ISBN 0-674-11649-6.
- ^ Гернси, Пол. "ЧТО ЕСТЬ ШИМПЫ?". Все о дикой природе. Получено 2013-04-22.
- ^ Ихобе Х (1992). "Наблюдения за мясоедом диких бонобо (Пан панискус) в Вамбе, Республика Заир ". Приматы. 33 (2): 247–250. Дои:10.1007 / BF02382754. S2CID 10063791.
- ^ Rafert, J .; Винеберг, Э. (1997). «Bonobo Nutrition - отношение диеты в неволе к диете» (PDF). Руководство по выращиванию бонобо. Американская ассоциация зоопарков и аквариумов. Архивировано из оригинал (PDF) на 2012-04-25.
- ^ Сурбек, М; Фаулер, А; Деймель, К; Хоманн, Г. (2008). "Свидетельства потребления бонобо древесных суточных приматов (Пан панискус)". Американский журнал приматологии. 71 (2): 171–4. Дои:10.1002 / ajp.20634. PMID 19058132. S2CID 32622605.
- ^ Surbeck M, Hohmann G; Хохманн (14 октября 2008 г.). «Охота на приматов бонобо в Луикотале, национальный парк Салонга». Текущая биология. 18 (19): R906–7. Дои:10.1016 / j.cub.2008.08.040. PMID 18957233. S2CID 6708310.
- ^ Кордейн Л., Итон С.Б., Себастьян А. и др. (Февраль 2005 г.). «Истоки и эволюция западной диеты: последствия для здоровья в 21 веке». Являюсь. J. Clin. Нутр. 81 (2): 341–54. Дои:10.1093 / ajcn.81.2.341. PMID 15699220.
- ^ Ulijaszek SJ (ноябрь 2002 г.). «Пищевое поведение человека в эволюционном экологическом контексте». Proc Nutr Soc. 61 (4): 517–26. Дои:10.1079 / PNS2002180. PMID 12691181.
- ^ Самое раннее сельское хозяйство в Америке В архиве 3 июня 2010 г. Wayback MachineРаннее выращивание ячменя В архиве 16 февраля 2007 г. Wayback MachineРаннее выращивание инжира В архиве 2 июня 2006 г. Wayback Machine, получено 19 февраля 2007 г.
- ^ Кребс-младший (сентябрь 2009 г.). «Обезьяна-гурман: эволюция и пищевые предпочтения человека». Являюсь. J. Clin. Нутр. 90 (3): 707С – 11С. Дои:10.3945 / ajcn.2009.27462B. PMID 19656837.
- ^ Холден С., Мейс Р. (октябрь 1997 г.). «Филогенетический анализ эволюции переваривания лактозы у взрослых». Гм. Биол. 69 (5): 605–28. PMID 9299882.
- ^ Фихтель, Клаудия (2012). «Хищничество». В Mitani, John C .; Позвони, Хосеп; Каппелер, Питер М .; Palombit, Ryne A .; Шелк, Джоан Б. (ред.). Эволюция обществ приматов. Издательство Чикагского университета. С. 169–84. ISBN 978-0-226-53172-4.
- ^ Boesch, C .; Бош, Х. (1990). «Использование инструментов и изготовление инструментов у диких шимпанзе». Folia Primatologica. 54 (1–2): 86–99. Дои:10.1159/000156428. PMID 2157651.
- ^ Westergaard, G.C .; Lundquist, A. L .; и другие. (1998). «Почему некоторые обезьяны капуцины (Cebus apella) используйте инструменты зондирования (а другие нет) ". Журнал сравнительной психологии. 112 (2): 207–211. Дои:10.1037/0735-7036.112.2.207. PMID 9642788.
- ^ де Ваал, Ф. Б. М .; Дэвис, Дж. М. (2003). «Когнитивная экология капуцинов: сотрудничество, основанное на прогнозируемой отдаче». Нейропсихология. 41 (2): 221–228. Дои:10.1016 / S0028-3932 (02) 00152-5. PMID 12459220. S2CID 8190458.
- ^ Paar, L.A .; Уинслоу, Дж. Т .; Hopkins, W. D .; де Ваал, Ф. Б. М. (2000). «Распознавание лицевых сигналов: индивидуальная дискриминация шимпанзе (Пан троглодиты) и макаки-резусы (Macaca mulatta)". Журнал сравнительной психологии. 114 (1): 47–60. Дои:10.1037/0735-7036.114.1.47. ЧВК 2018744. PMID 10739311.
- ^ Бирн, Ричард; Корп, Надя (2004). «Размер неокортекса предсказывает скорость обмана у приматов». Труды Лондонского королевского общества. Серия B: Биологические науки. 271 (1549): 1693–1699. Дои:10.1098 / rspb.2004.2780. ЧВК 1691785. PMID 15306289.
- ^ Paar, L.A .; де Ваал, Ф. Б. М. (1999). «Визуальное распознавание родства у шимпанзе». Природа. 399 (6737): 647–648. Bibcode:1999Натура.399..647P. Дои:10.1038/21345. PMID 10385114. S2CID 4424086.
- ^ Fujita, K .; Watanabe, K .; Widarto, T. H .; Сурёброто, Б. (1997). «Дискриминация макак макаками: случай видов сулавеси». Приматы. 38 (3): 233–245. Дои:10.1007 / BF02381612. S2CID 21042762.
- ^ Колл, Дж. (2001). «Постоянство объекта у орангутанов (Pongo pygmaeus), шимпанзе (Пан троглодиты), и дети (Homo sapiens)". Журнал сравнительной психологии. 115 (2): 159–171. Дои:10.1037/0735-7036.115.2.159. PMID 11459163.
- ^ Itakura, S .; Танака, М. (июнь 1998 г.). "Использование шимпанзе подсказок экспериментатора во время заданий по выбору предметов (Пан троглодиты), орангутанг (Pongo pygmaeus) и человеческие младенцы (Homo sapiens)". Журнал сравнительной психологии. 112 (2): 119–126. Дои:10.1037/0735-7036.112.2.119. PMID 9642782.
- ^ Gouteux, S .; Thinus-Blanc, C .; Воклер, Дж. (2001). «Обезьяны-резусы используют геометрическую и негеометрическую информацию во время задачи переориентации» (PDF). Журнал экспериментальной психологии: Общие. 130 (3): 505–519. Дои:10.1037/0096-3445.130.3.505.
- ^ Томаселло, М. и Колл, Дж. (1997). Познание приматов. Oxford University Press, США. ISBN 978-0-19-510624-4.
- ^ а б c Deaner, R. O .; van Schaik, C.P .; Джонсон, В. Э. (2006). «Есть ли у некоторых таксонов лучшее общее познание предметной области, чем у других? Метаанализ исследований нечеловеческих приматов» (PDF). Эволюционная психология. 4: 149–196. Дои:10.1177/147470490600400114. S2CID 16702785. Архивировано из оригинал (PDF) на 2011-07-28. Получено 2011-07-04.
- ^ а б c Читатель, С. М .; Hager, Y .; Лаланд, К. Н. (2011). «Эволюция общего и культурного интеллекта приматов» (PDF). Философские труды Королевского общества B. 366 (1567): 1017–1027. Дои:10.1098 / rstb.2010.0342. ЧВК 3049098. PMID 21357224. Архивировано из оригинал (PDF) на 2011-10-03. Получено 2011-07-04.
- ^ а б «Использование инструментов, охота и другие открытия». Институт Джейн Гудолл. Получено 2013-08-01.
- ^ "Бонобо". ApeTag. 2010. Архивировано с оригинал на 2013-11-02. Получено 2013-08-03.
- ^ Грубер, Т .; Clay, Z .; Цубербюлер, К. (2010). "Сравнение использования инструментов бонобо и шимпанзе: свидетельство предвзятости самок в Сковорода родословная " (PDF). Поведение животных. 80 (6): 1023–1033. Дои:10.1016 / j.anbehav.2010.09.005. S2CID 14923158.
- ^ Бауэр, Б. (18 апреля 2011 г.). «Орангутаны ловят рыбу с помощью простых инструментов». Проводной. Получено 2013-08-05.
- ^ Брейер, Т .; Ndoundou-Hockemba, M .; Фишлок, В. (2005). «Первое наблюдение использования орудия у диких горилл». PLOS Биология. 3 (11): e380. Дои:10.1371 / journal.pbio.0030380. ЧВК 1236726. PMID 16187795.
- ^ Fragaszy, D .; Изар, П .; Visalberghi, E .; Ottoni, E.B .; де Оливейра, М. (2004). «Дикие обезьяны капуцины (Cebus libidinosus) используйте наковальни и инструменты для забивания камня ». Американский журнал приматологии. 64 (4): 359–366. Дои:10.1002 / ajp.20085. PMID 15580579. S2CID 16222308.
- ^ Gumert, M.D .; Kluck, M .; Малаивиджитнонд, С. (2009). «Физические характеристики и образцы использования каменного топора и молотков, используемых длиннохвостыми макаками в районе Андаманского моря в Таиланде». Американский журнал приматологии. 71 (7): 594–608. Дои:10.1002 / ajp.20694. PMID 19405083. S2CID 22384150.
- ^ Гамильтон, W.J .; Buskirk, R.E .; Бускерк, W.H. (1975). «Защитное забивание камнями павианов». Природа. 256 (5517): 488–489. Bibcode:1975Натура.256..488H. Дои:10.1038 / 256488a0. S2CID 4149862.
- ^ Fichtel, C .; Каппелер, П. М. (2010). «Глава 19: Человеческие универсалии и симплезиоморфии приматов: установление исходной линии лемура». В Kappeler, P. M .; Силк, Дж. Б. (ред.). Не упускайте из виду: прослеживая истоки человеческих универсалий. Springer. ISBN 978-3-642-02724-6.
- ^ Сугияма, Ю. (1995). «Инструменты для питья диких шимпанзе в Босу». Американский журнал приматологии. 37 (1): 263–269. Дои:10.1002 / ajp.1350370308. PMID 31936951. S2CID 86473603.
- ^ «Суматранские орангутаны». OrangutanIslands.com. Архивировано из оригинал в 2013-11-26. Получено 2013-08-02.
- ^ van Schaik, C .; Fox, E .; Ситомпул, А. (1996). «Изготовление и использование инструментов в диких суматранских орангутанах». Naturwissenschaften. 83 (4): 186–188. Bibcode:1996NW ..... 83..186V. Дои:10.1007 / BF01143062. PMID 8643126. S2CID 27180148.
- ^ Гилл, Виктория (22 июля 2011 г.). "Обезьяна мандрил делает инструмент для педикюра". BBC. Получено 2013-08-11.
- ^ Ванкатова, М. (2008). «Гориллы и инструменты - Часть I». Получено 2013-08-04.
- ^ а б Сведелл, Лариса (2013). «Эволюция обществ приматов. Под редакцией Джона Митани, Джозепа. Вызов, Питер Каппелер, Райн Паломбит и Джоан Силк. Xiv + 730 стр. Чикаго, Иллинойс: University of Chicago Press. 2012. 145 долларов США (ткань), 50 долларов США (бумага), 7-40 долларов США (электронная книга) ". Американский журнал биологии человека. 25 (4): 571–572. Дои:10.1002 / ajhb.22412. Получено 2020-10-29.
- ^ Райт, Патрисия К. (1 апреля 1990 г.). «Образцы отцовской заботы у приматов». Международный журнал приматологии. 11 (2): 89–102. Дои:10.1007 / BF02192783. ISSN 1573-8604. S2CID 35420972.
- ^ Сведелл, Лариса (2013). "Эволюция обществ приматов. Под редакцией Джона Митани, Джозепа Вызов, Питер Каппелер, Райн Паломбит и Джоан Силк. Xiv + 730 стр. Чикаго, Иллинойс: University of Chicago Press. 2012. 145 долларов США (ткань), 50 долларов США (бумага), 7-40 долларов США (электронная книга) ". Американский журнал биологии человека. 25 (4): 571–572. Дои:10.1002 / ajhb.22412. Получено 2020-10-29.
- ^ Сведелл, Лариса (2013). «Эволюция обществ приматов. Под редакцией Джона Митани, Джозепа. Вызов, Питер Каппелер, Райн Паломбит и Джоан Силк. Xiv + 730 стр. Чикаго, Иллинойс: University of Chicago Press. 2012. 145 долларов США (ткань), 50 долларов США (бумага), 7-40 долларов США (электронная книга) ". Американский журнал биологии человека. 25 (4): 571–572. Дои:10.1002 / ajhb.22412. Получено 2020-10-29.
- ^ Сведелл, Лариса (2013). «Эволюция обществ приматов. Под редакцией Джона Митани, Джозепа. Вызов, Питер Каппелер, Райн Паломбит и Джоан Силк. Xiv + 730 стр. Чикаго, Иллинойс: University of Chicago Press. 2012. 145 долларов США (ткань), 50 долларов США (бумага), 7-40 долларов США (электронная книга) ". Американский журнал биологии человека. 25 (4): 571–572. Дои:10.1002 / ajhb.22412. Получено 2020-10-29.
- ^ Вайтт, Корри; Литтл, Энтони С .; Вулфенсон, Сара; Честное слово, Пол; Браун, Энтони П .; Бьюкенен-Смит, Ханна М .; Перретт, Дэвид И. (07.11.2003). «Данные, полученные на макаках-резус, свидетельствуют о том, что окраска самцов играет роль в выборе самки приматов». Труды Лондонского королевского общества. Серия B: Биологические науки. 270 (Suppl_2): S144 – S146. Дои:10.1098 / рсбл.2003.0065. ЧВК 1809959. PMID 14667364.
- ^ Вайтт, Корри; Литтл, Энтони С. (01.02.2006). «Предпочтения симметрии конкретной формы лица у Macaca mulatta». Международный журнал приматологии. 27 (1): 133–145. Дои:10.1007 / s10764-005-9015-у. ISSN 1573-8604. S2CID 20176410.
- ^ а б c Эстрада, Алехандро; Гарбер, Пол А .; Rylands, Энтони Б .; Роос, Кристиан; Фернандес-Дуке, Эдуардо; Фиоре, Энтони Ди; Некарис, К. Анн-Изола; Ниджман, Винсент; Хейманн, Экхард В. (01.01.2017). «Надвигающийся кризис вымирания приматов в мире: почему приматы имеют значение». Достижения науки. 3 (1): e1600946. Bibcode:2017SciA .... 3E0946E. Дои:10.1126 / sciadv.1600946. ISSN 2375-2548. ЧВК 5242557. PMID 28116351.
- ^ Сведелл, Лариса (2013). «Эволюция обществ приматов. Под редакцией Джона Митани, Джозепа. Вызов, Питер Каппелер, Райн Паломбит и Джоан Силк. Xiv + 730 стр. Чикаго, Иллинойс: University of Chicago Press. 2012. 145 долларов США (ткань), 50 долларов США (бумага), 7-40 долларов США (электронная книга) ". Американский журнал биологии человека. 25 (4): 571–572. Дои:10.1002 / ajhb.22412. Получено 2020-10-29.
- ^ Mays, Herman L .; Хилл, Джеффри Э. (октябрь 2004 г.). «Выбор партнера: хорошие гены против генов, которые подходят». Тенденции в экологии и эволюции. 19 (10): 554–559. Дои:10.1016 / j.tree.2004.07.018. ISSN 0169-5347. PMID 16701321.
- ^ Кнапп, Лесли А .; Робсон, Джули; Уотерхаус, Джон С. (2006). «Обонятельные сигналы и MHC: обзор и тематическое исследование у лемура катта». Американский журнал приматологии. 68 (6): 568–584. Дои:10.1002 / ajp.20253. ISSN 1098-2345. PMID 16715507. S2CID 45753542.
- ^ Кедди, Энн С. (1986). «Выбор самки у верветок (Cercopithecus aethiops sabaeus)». Американский журнал приматологии. 10 (2): 125–134. Дои:10.1002 / ajp.1350100204. ISSN 1098-2345. PMID 31979492. S2CID 83590177.
- ^ Kotiaho, J. S .; Пууртинен, М. (2007). «Выбор партнера для косвенных генетических выгод: исследование существующей парадигмы». Функциональная экология. 21 (4): 638–644. Дои:10.1111 / j.1365-2435.2007.01286.x. ISSN 1365-2435.
- ^ Хаффман, Майкл А. (1987). «Вторжение супруга и выбор самки у японских макак (Macaca fuscata)». Этология. 75 (3): 221–234. Дои:10.1111 / j.1439-0310.1987.tb00655.x. ISSN 1439-0310.
- ^ Cowlishaw, G .; Клаттон-Брок, Т. (2009). «Приматы». В Макдональде, Д. (ред.). Принстонская энциклопедия млекопитающих. Издательство Принстонского и Оксфордского университетов. С. 270–280. ISBN 978-0-691-14069-8.
- ^ Рид, К .; Флигл, Дж. (15 августа 1995 г.). «Географический и климатический контроль разнообразия приматов». Труды Национальной академии наук Соединенных Штатов Америки. 92 (17): 7874–7876. Bibcode:1995PNAS ... 92.7874R. Дои:10.1073 / пнас.92.17.7874. ЧВК 41248. PMID 7644506.
- ^ Чепмен, С. и Руссо, С. (2007). "Распространение семян приматов". В Campbell, C.J .; Fuentes, A .; MacKinnon, K. C .; Пангер М. и Бердер С. К. (ред.). Приматы в перспективе. Издательство Оксфордского университета. п. 510. ISBN 978-0-19-517133-4.
- ^ Long, Y. C .; Киркпатрик, Р. С.; Чжун, Т .; Сяо, Л. (апрель 1994 г.). "Отчет о распространении, популяции и экологии курносой обезьяны провинции Юньнань (Ринопитек bieti)". Приматы. 35 (2): 241–250. Дои:10.1007 / BF02382060. S2CID 23931368.
- ^ Шаллер, Г. Б. (1963). Горная горилла: экология и поведение. Чикаго: University Chicago Press. ISBN 978-0-226-73635-8.
- ^ Штаммбах, Э. (1987). "Пустыня, лес и горные бабуины: многоуровневые общества". В Smuts, B; Чейни, Д.; Seyfarth, R; Wrangham, R; Struhsaker, T. (ред.). Общества приматов. Издательство Чикагского университета. С. 112–120. ISBN 978-0226767161.
- ^ Кемп, Э. (2009). «Модели водопользования у приматов». Folia Primatologica. 80 (4): 275–294. Дои:10.1159/000252586. PMID 19864919. S2CID 5108827.
- ^ Вулф, Л. Д. и Фуэнтес, А. (2007). «Этноприматология». В Campbell, C.J .; Fuentes, A .; MacKinnon, K. C .; Пангер М. и Бердер С. К. (ред.). Приматы в перспективе. Издательство Оксфордского университета. п. 692. ISBN 978-0-19-517133-4.
- ^ Ренквист Д. М. и Уитни Р. А. (1987). «Зоонозы, полученные от домашних приматов». Ветеринарные клиники Северной Америки: практика мелких животных. 17 (1): 219–240. Дои:10.1016 / s0195-5616 (87) 50614-3. PMID 3551307. Получено 2008-08-11.
- ^ «Всеобщая декларация прав человека». Объединенные Нации. 1948. Архивировано с оригинал на 2014-12-08. Получено 2008-12-02.
- ^ а б Кавальери П. и Сингер П. "Декларация о больших обезьянах". Великая обезьяна. Архивировано из оригинал на 2008-08-20. Получено 2008-06-16.
- ^ Глендиннинг, Л. (26 июня 2008 г.). «Испанский парламент одобряет« права человека »для обезьян». Хранитель. Получено 2008-11-10.
- ^ Певица П. (18 июля 2008 г.). «Из человекообразных обезьян и людей». Хранитель. Получено 2008-11-10.
- ^ Мотт, М. (16 сентября 2003 г.). "Опасности содержания обезьян в качестве домашних животных". Национальная география. Архивировано из оригинал на 2013-06-08. Получено 2013-02-06.
- ^ а б Уоркман, К. (июнь 2004 г.). «Сохранение приматов во Вьетнаме: к целостному экологическому повествованию». Американский антрополог. 106 (2): 346–352. Дои:10.1525 / aa.2004.106.2.346.
- ^ "Новости IPPL: Торговля обезьянами в США". Международная лига защиты приматов. 2003. Архивировано с оригинал на 2008-07-25. Получено 2008-08-04.
- ^ Бушнелл, Д. (1958). "Начало исследований в области космической биологии в Центре разработки ракет ВВС, 1946–1952 гг.". История исследований в области космической биологии и биодинамики. НАСА. Получено 2008-08-18.
- ^ Блюменталь, Д. (1987-06-17). "Обезьяны как помощники парализованным в домашних условиях". Нью-Йорк Таймс. Получено 2008-10-08.
- ^ Ньюман, Джеймс Л. (2013). Встреча с гориллами: хроника открытий, эксплуатации, понимания и выживания. Плимут, Соединенное Королевство: Роуман и Литтлфилд. п.173. ISBN 978-1-4422-1957-1.
- ^ а б «Поставки и использование приматов в ЕС». Европейская ассоциация биомедицинских исследований. 1996. Архивировано с оригинал на 2012-01-17. Получено 2008-08-18.
- ^ Chen, F.C .; Ли, В. Х. (февраль 2001 г.). «Геномные расхождения между людьми и другими гоминоидами и эффективный размер популяции общего предка людей и шимпанзе». Американский журнал генетики человека. 68 (2): 444–456. Дои:10.1086/318206. ЧВК 1235277. PMID 11170892.
- ^ Конли, К. М .; Hoffeld, E. H .; Стивенс, М. Л. (2004). «Демографический анализ исследований приматов в Соединенных Штатах». Альтернативы лабораторным животным. 32 (Sup 1): 315–322. Дои:10.1177 / 026119290403201s52. PMID 23577480. S2CID 6743973.
- ^ Представлено парламенту министром внутренних дел по приказу Ее Величества (июль 2006 г.). Статистика научных процедур на живых животных: Великобритания 2005 г. (PDF). Канцелярский офис. ISBN 0-10-168772-9. Получено 2008-06-16.
- ^ «Нечеловеческие приматы: животные для исследований». Информационный центр защиты животных. Министерство сельского хозяйства США. Архивировано из оригинал на 2008-10-03. Получено 2008-07-14.
- ^ «Директива 86/609». Европейская коалиция за прекращение экспериментов на животных. Архивировано из оригинал на 2008-09-29. Получено 2008-10-08.
- ^ IFAW (2005). Рождены дикими животными: приматы - не домашние животные (PDF). Международный фонд защиты животных. Архивировано из оригинал (PDF) на 2011-07-26. Получено 2011-02-26.
- ^ СИТЕС (2010-10-14). «Приложения I, II и III». Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения. Получено 2012-04-02.
- ^ Грабб, П. (1998). «Сьерра-леонская обезьяна едет». Млекопитающие Ганы, Сьерра-Леоне и Гамбии. Сент-Айвс: Трендрин. С. 214–219. ISBN 0-9512562-4-6.
- ^ а б Chapman, C.A .; Перес, К. А. (2001). «Сохранение приматов в новом тысячелетии: роль ученых». Эволюционная антропология. 10 (1): 16–33. Дои:10.1002 / 1520-6505 (2001) 10: 1 <16 :: AID-EVAN1010> 3.0.CO; 2-O.
- ^ а б Миттермайер, Р. А. и Чейни, Д. Л. (1987). «Сохранение приматов и среды их обитания». В Smuts, B. B .; Cheney, D. L .; Seyfarth, R.M .; Wrangham, R. W. & Struhsaker, T. T. (ред.). Общества приматов. Чикаго: Издательство Чикагского университета. С. 477–490.
- ^ а б Саутвик, К. Х. и Сиддики, М. Ф. (2001). «Статус, сохранение и управление приматами в Индии» (PDF). Бюллетень Envis: дикая природа и охраняемые территории. 1 (1): 81–91. Архивировано из оригинал (PDF) на 2008-10-01. Получено 2008-08-04.
- ^ а б c d Коулишоу, Г. и Данбар, Р. (2000). Биология сохранения приматов. Чикаго: Издательство Чикагского университета. ISBN 978-0-226-11637-2.
- ^ Van Schaik, C.P .; Монк, К. А .; Робертсон, Дж. М. Ю. (2001). «Резкое сокращение численности орангутанов в экосистеме Лейзер на севере Суматры». Орикс. 35 (1): 14–25. Дои:10.1046 / j.1365-3008.2001.00150.x.
- ^ Purvis, A .; Gittleman, J. L .; Cowlishaw, G .; Мейс, Г. М. (2000). «Прогнозирование риска исчезновения исчезающих видов». Труды Королевского общества B. 267 (1456): 1947–1952. Дои:10.1098 / rspb.2000.1234. ЧВК 1690772. PMID 11075706.
- ^ а б c Fa, J. E .; Juste, J .; Perez de Val, J .; Кастровьехо, Дж. (1995). «Влияние рыночной охоты на виды млекопитающих в Экваториальной Гвинее». Биология сохранения. 9 (5): 1107–1115. Дои:10.1046 / j.1523-1739.1995.9051107.x. HDL:10261/49187.
- ^ Хилл, К. М. (1997). «Рейды диких позвоночных: взгляд фермера в сельскохозяйственном сообществе в западной Уганде». Международный журнал борьбы с вредителями. 43 (1): 77–84. Дои:10.1080/096708797229022.
- ^ Хилл, К. М. (2002). «Сохранение приматов и местные сообщества: этические вопросы и дебаты». Американский антрополог. 104 (4): 1184–1194. Дои:10.1525 / aa.2002.104.4.1184.
- ^ Чоудхури, А. (2001). «Приматы в Северо-Восточной Индии: обзор их распространения и статуса сохранения» (PDF). Бюллетень Envis: дикая природа и охраняемые территории. 1 (1): 92–101. Архивировано из оригинал (PDF) на 2008-10-01. Получено 2008-08-04.
- ^ Kumara, H.N .; Сингх М. (октябрь 2004 г.). "Распространение и численность приматов в тропических лесах Западных Гат, Карнатака, Индия и сохранение Макака силенус". Международный журнал приматологии. 25 (5): 1001–1018. Дои:10.1023 / B: IJOP.0000043348.06255.7f. S2CID 30384142.
- ^ Ниджман, В. (2004). "Сохранение яванского гиббона Гилобаты молоха: оценки численности населения, локальное вымирание и приоритеты сохранения " (PDF). Зоологический бюллетень Raffles. 52 (1): 271–280. Архивировано из оригинал (PDF) на 2008-09-10. Получено 2008-08-04.
- ^ O'Brien, T. G .; Kinnaird, M. F .; Nurcahyo, A .; Iqbal, M .; Русманто, М. (апрель 2004 г.). «Изобилие и распространение симпатрических гиббонов в тропических лесах Суматры, находящихся под угрозой исчезновения». Международный журнал приматологии. 25 (2): 267–284. Дои:10.1023 / B: IJOP.0000019152.83883.1c. S2CID 32472118.
- ^ Эстрада, А .; Coates-Estrada, R .; Меритт, Д. (сентябрь 1994 г.). «Нелетающие млекопитающие и изменения ландшафта в районе тропических лесов Лос-Тустлас, Мексика». Экография. 17 (3): 229–241. Дои:10.1111 / j.1600-0587.1994.tb00098.x.
- ^ Марш, Л. К. (2003). «Природа раздробленности». В Марше, Л. К. (ред.). Приматы в фрагментах: экология и сохранение. Нью-Йорк: Kluwer Academic /Издатели Пленума. С. 1–10. ISBN 0-306-47696-7.
- ^ Тернер, И. М. (1996). «Гибель видов во фрагментах влажных тропических лесов: обзор свидетельств». Журнал прикладной экологии. 33 (2): 200–209. Дои:10.2307/2404743. JSTOR 2404743.
- ^ Кьярелло, А.Г. (2003). «Приматы бразильского атлантического леса: влияние фрагментации леса на выживание». В Марше, Л. К. (ред.). Приматы в фрагментах: экология и сохранение. Нью-Йорк: Kluwer Academic / Plenum Publishers. С. 99–121. ISBN 978-0-306-47696-9.
- ^ Поуп, Т. (1996). «Социоэкология, фрагментация популяции и закономерности генетической потери у находящихся под угрозой исчезновения приматов». В Avise, J .; Хэмрик, Дж. (Ред.). Сохранение генетики: истории болезни природы. Norwell: Kluwer Academic Publishers. С. 119–159. ISBN 978-0-412-05581-2.
- ^ Миттермайер, Р.А.; Wallis, J .; Rylands, A.B .; Ganzhorn, J.U .; Oates, J.F .; Уильямсон, E.A .; Palacios, E .; Heymann, E.W .; Kierulff, M.C.M .; Yongcheng, L .; Supriatna, J .; Roos, C .; Уокер, S .; Cortés-Ortiz, L .; Швитцер, К., ред. (2009). Приматы в опасности: 25 самых угрожаемых приматов в мире, 2008–2010 гг. (PDF). В иллюстрациях С.Д. Нэш. Арлингтон, Вирджиния: Группа специалистов по приматам МСОП / SSC (PSG), Международное приматологическое общество (IPS) и Conservation International (CI). С. 23–26. ISBN 978-1-934151-34-1.
- ^ Oates, J. F .; Abedi-Lartey, M .; McGraw, W. S .; Struhsaker, T. T .; Уайтсайдс, Г. Х. (октябрь 2000 г.). «Вымирание западноафриканской красной обезьяны колобус». Биология сохранения. 14 (5): 1526–1532. Дои:10.1046 / j.1523-1739.2000.99230.x.
- ^ МакГроу, В. С. (июнь 2005 г.). «Обновленная информация о поисках красной обезьяны колобус мисс Уолдрон». Международный журнал приматологии. 26 (3): 605–619. Дои:10.1007 / s10764-005-4368-9. S2CID 861418.
Цитированная литература
- Бентон, Майкл Дж. (2005). «Глава 3: Эволюция приматов». Палеонтология позвоночных. Вили-Блэквелл. ISBN 978-0-632-05637-8. Получено 2011-07-10.
- Картмилл, М. (2010). "Классификация приматов и разнообразие". In Platt, M .; Ghazanfar, A (ред.). Нейроэтология приматов. Издательство Оксфордского университета. С. 10–30. ISBN 978-0-19-532659-8.
- Хартвиг, В. (2011). «Глава 3: Эволюция приматов». В Campbell, C.J .; Fuentes, A .; MacKinnon, K. C .; Bearder, S.K .; Штумпф, Р. М. (ред.). Приматы в перспективе (2-е изд.). Издательство Оксфордского университета. С. 19–31. ISBN 978-0-19-539043-8.
- Szalay, F.S .; Делсон, Э. (1980). Эволюционная история приматов. Академическая пресса. ISBN 978-0126801507. OCLC 893740473.
дальнейшее чтение
| Библиотечные ресурсы о Приматы |
- Дэвид Дж. Чиверс; Бернард А. Вуд; Алан Билсборо, ред. (1984). Приобретение и переработка пищи у приматов. Нью-Йорк и Лондон: Пленум Пресс. ISBN 0-306-41701-4.
внешняя ссылка
- Примас в Керли
- Информационная сеть примаса
- Приматы в Сеть разнообразия животных
- Научно-исследовательский институт приматов, Киотский университет
- Цитоархитектурные атласы мозга приматов высокого разрешения
- EUPRIM-Net: Европейская сеть приматов
- PrimateImages: Коллекция естественной истории
- Интерактивные представления скелетов различных приматов на eSkeletons.org (связанных с Техасский университет в Остине )
- Веб-проект "Древо жизни"