Эволюция четвероногих - Evolution of tetrapods
| Часть серии по |
| Палеонтология |
|---|
 |
История палеонтологии |
Отрасли палеонтологии |
| Палеонтологический портал Категория |
В эволюция четвероногих началось около 400 миллионов лет назад в Девонский период с самого раннего четвероногие развился из рыба с лопастными плавниками.[1] Четвероногие (под апоморфия -основанное определение, используемое на этой странице), относятся к категории животных в биологический суперкласс Тетрапода, куда входят все живые и вымершие амфибии, рептилии, птицы, и млекопитающие. В то время как большинство видов сегодня являются наземными, мало доказательств поддерживает идею о том, что какие-либо из самых ранних четвероногих могли передвигаться по суше, поскольку их конечности не могли удерживать свое тело от земли, а известные следы не указывают на то, что они волочили живот. Предположительно, следы оставили животные, идущие по дну неглубоких водоемов.[2] Конкретные водные предки четвероногих и процесс колонизации суши остаются неясными и являются областями активных исследований и дискуссий среди палеонтологи в настоящий момент.
Большинство земноводных сегодня остаются полуводными, проживая первую стадию своей жизни как рыбы. головастики. Несколько групп четвероногих, таких как змеи и китообразные, потеряли некоторые или все конечности. Кроме того, многие четвероногие вернулись к частично водной или полностью водной жизни на протяжении всей истории группы (современные примеры полностью водных четвероногих включают китообразных и сирены ). Первое возвращение к водному образу жизни могло произойти еще в Каменноугольный период[3] тогда как другие возвраты произошли совсем недавно, Кайнозойский, как у китообразных, ластоногие,[4] и несколько современные амфибии.[5]
Переход от плана тела для дыхания и навигации в воде к плану тела, позволяющему животному перемещаться по суше, является одним из наиболее глубоких известных эволюционных изменений.[6] Это также один из наиболее понятных, во многом благодаря ряду важных переходное ископаемое находки конца 20 века в сочетании с улучшенным филогенетическим анализом.[1]
Источник
Эволюция рыбы
Девонский период традиционно известен как «Эпоха рыб», отмечая диверсификацию многочисленных вымерших и современных основных групп рыб.[7] Среди них были ранние костлявые рыбы, которые в начале периода разнообразились и распространились в пресноводных и солоноватых средах. Ранние типы напоминали свои хрящевые предки во многих особенностях анатомии, включая акульоподобный хвостовой плавник, спиральную кишку, большой грудные плавники усиленный спереди скелетными элементами и в значительной степени неокостеневшим осевой скелет.[8]
Однако у них были определенные черты, отделяющие их от хрящевых рыб, черты, которые стали ключевыми в эволюции наземных форм. За исключением пары дыхальца, то жабры не открывались только внешнему виду, как акулы; скорее, они были заключены в жаберную камеру, укрепленную мембранные кости и покрыт костлявым крышка, с единственным выходом наружу. В кость клейтрума, образующие задний край жаберной камеры, также служат якорем для грудных плавников. У хрящевых рыб такого крепления для грудных плавников нет. Это позволило создать подвижный сустав в основании плавников у ранних костистых рыб, а позже он будет функционировать в качестве несущей конструкции у четвероногих. В составе общей брони ромбовидный Космин весы, то череп была полная обложка кожная кость, составляя крыша черепа над акулоподобным хрящом внутренний череп. Важно отметить, что у них также была пара вентральные парные легкие,[9] особенность, отсутствующая у акул и скатов.
Предполагалось, что рыбы в значительной степени эволюционировали вокруг рифы, но с момента своего возникновения около 480 миллионов лет назад они жили в прибрежных средах, таких как приливные зоны или постоянно мелководные лагуны, и не начали распространяться в другие биотопы раньше, чем 60 миллионов лет спустя. Некоторые приспособились к более глубокой воде, в то время как твердые и массивные формы остались там, где они были, или мигрировали в пресную воду.[10][11] Увеличение первичной продуктивности на суше в конце девона изменило пресноводные экосистемы. Когда питательные вещества из растений попадали в озера и реки, они поглощались микроорганизмами, которые, в свою очередь, поедались беспозвоночными, которые служили пищей для позвоночных. Некоторые рыбы тоже стали детритофаги.[12] Ранние четвероногие развили терпимость к средам с разной соленостью, таким как эстуарии или дельты.[13]
Легкие раньше земли
Легкие / плавательный пузырь возникли как разрастание кишечника, образуя наполненный газом пузырь над пищеварительной системой. В своей примитивной форме воздушный пузырь был открыт для пищеварительный тракт, условие называется физостом и до сих пор встречается во многих рыбах.[14] Основная функция не совсем определена. Одно соображение плавучесть. Тяжелая чешуйчатая броня ранних костистых рыб, несомненно, утяжеляла животных. У хрящевых рыб, лишенных плавательного пузыря, акулам открытого моря необходимо постоянно плавать, чтобы не погрузиться на глубину, а грудные плавники обеспечивают поднимать.[15] Другой фактор - потребление кислорода. Кислород в окружающей среде в раннем девоне был относительно низким, возможно, около половины современных значений.[16] На единицу объема в воздухе гораздо больше кислорода, чем в воде, а позвоночные - активные животные с высокими потребностями в энергии по сравнению с беспозвоночными аналогичного размера.[17][18] В девоне наблюдалось повышение уровня кислорода, что открыло новые экологические ниши, позволив группам, которые могли использовать дополнительный кислород, развиться в активных, крупных животных.[16] В частности, в местах обитания на тропических болотах атмосферный кислород гораздо более стабилен и, возможно, заставил полагаться на легкие, а не на жабры для поглощения первичного кислорода.[19][20] В конце концов, плавучесть и дыхание, возможно, были важны, и некоторые современные физиостомные рыбы действительно используют свой мочевой пузырь для обоих.
Для газообмена легким требуется кровоснабжение. У хрящевых рыб и костистые кости, то сердце лежит низко в теле и качает кровь вперед через брюшная аорта, который разделяется на серию парных дуг аорты, каждая из которых соответствует жаберная дуга.[21] Затем дуги аорты сливаются над жабрами, образуя дорсальная аорта снабжение организма насыщенной кислородом кровью. В двоякодышащие, Bowfin и бихиры кровоснабжение плавательного пузыря осуществляется парными легочные артерии ответвляется от задней (6-й) дуги аорты.[22] Такая же основная картина наблюдается у двоякодышащих рыб. Протоптерус и в земных саламандры, и, вероятно, это был образец, обнаруженный у непосредственных предков четвероногих, а также у первых четвероногих.[23] У большинства других костных рыб плавательный пузырь снабжается кровью через спинную аорту.[22]
Дыхание
Чтобы легкие могли производить газообмен, в них сначала должен быть газ. У современных четвероногих от ранних предков сохранились три важных механизма дыхания, первым из которых является СО.2/ H + система обнаружения. При дыхании современных четвероногих импульс к вдоху вызывается накоплением CO.2 в кровотоке и не недостаток O2.[24] Аналогичный СО2/ H + система обнаружения встречается во всех Osteichthyes, откуда следует, что последний общий предок Все Остейхти нуждались в такой системе обнаружения.[24][25] Второй механизм дыхания - это поверхностно-активное вещество система в легких для облегчения газообмена. Это также встречается во всех Osteichthyes, даже в тех, которые почти полностью водные.[26][27] Высококонсервативный характер этой системы предполагает, что даже водные Остейхти нуждаются в системе поверхностно-активных веществ, что может показаться странным, поскольку под водой нет газа. Третий механизм дыхания - это собственное движение дыхания. Этот механизм предшествует последнему общему предку Osteichthyes, что можно наблюдать у Lampetra camtshatica, то сестра клада Остейхтиесу. У миног этот механизм принимает форму «кашля», когда минога трясет своим телом, позволяя воде течь по жабрам. Когда CO2 уровни в крови миноги становятся слишком высокими, на центральный генератор сигналов посылается сигнал, который заставляет миногу "кашлять" и пропускать CO2 покинуть свое тело.[28][29] Эта связь между CO2 Система обнаружения и центральный генератор паттернов очень похожи на связь между этими двумя системами у четвероногих, что подразумевает гомологию.
Наружные и внутренние ноздри
В ноздри у большинства костистых рыб отличаются от таковых у четвероногих. Обычно у костистых рыб четыре ноздри (носовые отверстия), по одному наре за другим с каждой стороны. По мере того как рыба плывет, вода течет в переднюю пару через обонятельная ткань, и через задние отверстия. Это верно не только для лучеплавниковой рыбы, но и для латимерия, рыба, входящая в Саркоптеригии, группа, в которую также входят четвероногие. Напротив, у четвероногих есть только одна пара ноздрей снаружи, но также есть пара внутренних ноздрей, называемых хоаны, позволяя им втягивать воздух через нос. Двоякодышащие также являются саркоптеригами с внутренними ноздрями, но они настолько отличаются от хоан четвероногих, что уже давно признаны самостоятельным развитием.[30]
В ХХ веке горячо обсуждалась эволюция ноздрей четвероногих. Внутренние ноздри могут быть одним набором внешних (обычно считается задней парой), которые мигрировали в рот, или внутренняя пара может быть недавно сформированной структурой. Однако, чтобы освободить место для миграции, две зубные кости верхней челюсти, верхняя челюсть и предчелюстная кость, пришлось бы отделиться, чтобы пропустить ноздрю, а затем воссоединиться; до недавнего времени не было свидетельств переходной стадии, когда две кости были разъединены. Такие доказательства теперь доступны: небольшая рыба с лопастными плавниками, называемая Кенихтис, найденный в Китае и датируемый примерно 395 миллионами лет, представляет собой эволюцию, «застрявшую в середине акта», с разделенными верхней и верхней челюстями, а также отверстием - зарождающейся хоаной - на губе между двумя костями.[31] Кенихтис более близок к четвероногим, чем латимерия,[32] который имеет только наружные ноздри; таким образом, он представляет собой промежуточную стадию в развитии состояния четвероногих. Однако причина эволюционного движения задней ноздри от носа к губе до конца не изучена.
На мелководье

Родственники Кенихтис вскоре обосновались в водных путях и солоноватых устьях и стали самой многочисленной костистой рыбой на протяжении всего Девона и большей части Каменноугольный. Основная анатомия группы хорошо известна благодаря очень детальной работе над Евстеноптерон к Эрик Ярвик во второй половине 20 века.[33] Кости крыша черепа были в целом подобны зубам ранних четвероногих, а эмаль зубов была вздута, как у лабиринтодонты. Парные плавники имели отчетливо выраженное строение с костями. гомологичный к плечевая кость, локтевая кость, и радиус в передних плавниках и в бедренная кость, большеберцовая кость, и малоберцовая кость в тазовых плавниках.[34]
Было несколько семей: Ризодонтида, Canowindridae, Elpistostegidae, Megalichthyidae, Остеолепидные и Tristichopteridae.[35] Большинство из них были рыбами в открытой воде, а некоторые выросли до очень больших размеров; взрослые экземпляры достигают нескольких метров в длину.[36] Ризодонтид Ризодус По оценкам, он вырос до 7 метров (23 фута), что делает его самой крупной из известных пресноводных рыб.[37]
Хотя большинство из них были рыбами в открытой воде, одна группа, Эльпистостегалицы, адаптированный к жизни на мелководье. Они развили плоские тела для движения на очень мелководье, а грудные и тазовые плавники взяли на себя роль основных органов движения. Большинство срединных плавников исчезло, осталось только протоцеркальный хвостовой плавник. Поскольку мелководье время от времени испытывало недостаток кислорода, способность дышать атмосферным воздухом с помощью плавательного пузыря становилась все более важной.[6] В дыхальце стали большими и заметными, что позволило этим рыбам втягивать воздух.
Морфология черепа
Четвероногие имеют свои корни в раннем Девонский рыба-тетраподоморф.[38] Примитивные четвероногие развились от остеолепидной тетраподоморфной рыбы с лопастными плавниками (саркоптеригиан-кроссоптеригиан), с двухлопастной мозг в уплощенном череп. Группа латимерии представляет собой морских саркоптеригов, которые никогда не приобретали эти мелководные приспособления. Саркоптериги, очевидно, взяли две разные линии происхождения и соответственно разделены на две основные группы: Актинистия (включая латимерии) и Рипидистия (которые включают вымершие линии рыб с лопастными плавниками, которые превратились в двоякодышащих и тетраподоморфов).
От плавников до ступней

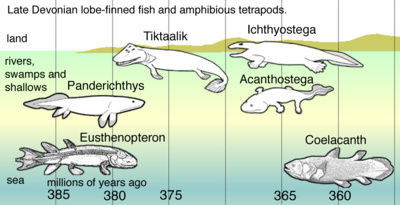
Самый старый из известных тетраподоморф - это Кенихтис из Китая, возраст которого составляет около 395 миллионов лет. Два самых ранних тетраподоморфа, датируемые 380 млн лет назад, были Гогонас и Пандерихтис.[39] Они имели хоаны и использовали свои плавники, чтобы перемещаться по приливным каналам и мелководью, забитой мертвыми ветками и гниющими растениями.[40] Их плавники могли быть использованы для прикрепления к растениям или тому подобному, когда они лежали в засаде для добычи. Универсальные четвероногие характеристики передней конечности которые наклоняются вперед от локоть и задние конечности, которые сгибаются назад от колено можно правдоподобно проследить до ранних четвероногих, живших на мелководье. Окаменелости тазовых костей из Тиктаалик показывает, если это типично для ранних четвероногих в целом, что задние придатки и движение с тазовым движением возникли в воде до земной адаптации.[41]
Еще один признак того, что лапы и другие черты четвероногих эволюционировали, когда животные были еще водными, - это способ их кормления. У них не было модификаций черепа и челюсти, которые позволяли бы глотать добычу на суше. Добычу можно было поймать на мелководье, у кромки воды или на суше, но ее нужно было есть в воде, где гидродинамические силы от расширения их ротовой полости заставляли пищу попадать в пищевод.[42]
Было высказано предположение, что эволюция конечностей четвероногих из плавников у рыб с лопастными плавниками связана с выражением HOXD13 ген или потеря белков актинодин 1 и актинодин 2, которые участвуют в развитии плавников рыб.[43][44] Моделирование роботов предполагает, что нервная система, необходимая для ходьбы, эволюционировала из нервов, управляющих плаванием, с использованием боковых движений. колебание тела с конечностями, в основном функционирующими как точки крепления и обеспечивающими ограниченную тягу.[45] Этот тип движения, а также изменения грудного пояса, аналогичные тем, которые наблюдаются в летописи окаменелостей, могут быть вызваны в бихиры подняв их из воды.[46]
Исследование 2012 года с использованием 3D-реконструкции Ихтиостега пришел к выводу, что он неспособен к типичному четвероногие походки. Конечности не могли двигаться попеременно, так как им не хватало необходимого диапазона вращательных движений. Кроме того, у задних конечностей отсутствовала мускулатура таза, необходимая для движения по земле с помощью задних конечностей. Их наиболее вероятный метод наземного передвижения - это синхронные «движения с костылями», аналогичные современным прыгуны.[47] (Просмотр нескольких видеозаписей "ходьбы" прыгуна показывает, что они двигаются, подтягивая себя вперед двумя грудными плавниками одновременно (левый и правый грудные плавники движутся одновременно, а не поочередно). Плавники выдвигаются вперед и опускаются; затем плечи вращаются. назад, продвигая корпус и волоча хвост в качестве третьей точки соприкосновения. Нет задних «конечностей» / плавников и нет значительного изгиба позвоночника.)
Обитатели болота
Вероятно, первые четвероногие развился в прибрежных и солоноватый морской средах, а также на мелководье и болотистый пресная вода среда обитания.[48] Раньше исследователи считали, что это конец девона. В 2010 году это убеждение было оспорено открытием старейших известных следов четвероногих, сохранившихся в морских отложениях южного побережья Лавразия, сейчас же Свентокшиские горы (Святой Крест) Польши. Они были сделаны во время Эйфельский этап в конце среднего девона. Следы, некоторые из которых содержат цифры, датируются примерно 395 миллионами лет назад - на 18 миллионов лет раньше, чем самые старые известные окаменелости тела четвероногих.[49] Кроме того, следы показывают, что животное было способно толкать руки и ноги вперед, тип движения, который был бы невозможен для рыб-тетраподоморфов, таких как Тиктаалик. По оценкам, животное, которое произвело следы, было до 2,5 метров (8,2 фута) в длину с подушечками лап до 26 сантиметров (10 дюймов) в ширину, хотя большинство следов всего 15 сантиметров (5,9 дюйма) в ширину.[50]
Новые находки предполагают, что первые четвероногие, возможно, жили как оппортунисты на приливных равнинах, питаясь морскими животными, которые были выброшены на берег или выброшены на берег приливом.[49] Однако в настоящее время рыба в значительном количестве выбрасывается на мель только в определенное время года, как в жена сезон нереста; такие высадки не могли обеспечить хищников значительным кормом. Нет никаких оснований предполагать, что девонские рыбы были менее благоразумными, чем современные.[51] По словам Мелины Хейл из Чикагского университета, не все древние следы обязательно созданы ранними четвероногими, но также могут быть созданы родственниками четвероногих, которые использовали свои мясистые придатки в аналогичных передвижениях на основе субстрата.[52][53]
Палеозойские четвероногие
Эта секция нужны дополнительные цитаты для проверка. (Ноябрь 2012 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |
Девонские четвероногие
Исследования Дженнифер А. Клак и ее коллеги показали, что самые ранние четвероногие животные, похожие на Акантостега, были полностью водными и совершенно непригодными для жизни на суше. Это контрастирует с более ранним представлением о том, что рыба сначала вторглась на сушу - либо в поисках добычи (как современные прыгуны ) или найти воду, когда пруд, в котором они жили, высох - а позже развились ноги, легкие и т. д.
К концу девона земля растения стабилизировала пресноводные среды обитания, что позволило первым водно-болотное угодье экосистемы развиваться со все более сложными пищевые полотна это открыло новые возможности. Пресноводные среды обитания были не единственными местами, где можно было найти воду, наполненную органическими веществами и густой растительностью, у самой кромки воды. В то время также существовали болотистые места обитания, такие как мелкие водно-болотные угодья, прибрежные лагуны и большие солоноватоводные речные дельты, и есть много оснований предполагать, что именно в такой среде развивались четвероногие. Ранние ископаемые четвероногие были обнаружены в морских отложениях, и поскольку окаменелости примитивных четвероногих в целом разбросаны по всему миру, они, должно быть, распространились по береговой линии - они не могли жить только в пресной воде.
Один анализ, проведенный Университетом Орегона, предполагает отсутствие доказательств теории «сокращающегося водоема» - переходные окаменелости не связаны с свидетельствами сокращения луж или прудов - и указывает на то, что такие животные, вероятно, не выжили бы в коротких переходах между истощенными водоемами.[54] Вместо этого новая теория предполагает, что протолёгкие и прото-конечности были полезными приспособлениями для преодоления условий окружающей среды во влажных лесных поймах.[55]
Девонские четвероногие прошли через два основных узких места во время так называемого Позднее девонское вымирание; один в конце Франский стадии, и один вдвое больше в конце следующего Фаменский сцена. Эти события вымирания привели к исчезновению примитивных четвероногих с рыбоподобными чертами, таких как Ichthyostega, и их основных водных родственников.[56] Когда четвероногие снова появляются в летописи окаменелостей после девонского вымирания, все взрослые формы полностью адаптированы к наземному существованию, а более поздние виды вторично адаптированы к водному образу жизни.[57]
Экскреция у четвероногих
Общий предок всех присутствующих гнатомы (челюстные позвоночные) жили в пресной воде, а позже мигрировали обратно в море.[нужна цитата ] Чтобы справиться с гораздо более высокой соленостью морской воды, они разработали способность поворачивать азот отходы аммиак в безобидный мочевина, сохраняя его в теле, чтобы дать крови то же самое осмолярность как морская вода, не отравляя организм. Это система, которая в настоящее время находится в хрящевые рыбы. Рыбы со скатными плавниками (Actinopterygii) позже вернулся в пресную воду и потерял эту способность, в то время как рыба с мясистыми плавниками (Sarcopterygii) сохранил его. Поскольку в крови рыб с лучевыми плавниками содержится больше соли, чем в пресной воде, они могут просто избавиться от аммиака через жабры. Когда они, наконец, снова вернулись в море, они не смогли восстановить свой старый трюк с превращением аммиака в мочевину, и вместо этого им пришлось развить солевые железы. Двудомные рыбы делают то же самое, когда они живут в воде, производя аммиак, а не мочевину, но когда вода высыхает и они вынуждены зарываться в грязь, они переключаются на производство мочевины. Подобно хрящевым рыбам, латимерия могут накапливать мочевину в своей крови, как и единственные известные земноводные, которые могут долгое время жить в соленой воде ( жаба Буфо маринус и лягушка Рана канкривора ). Это черты, которые они унаследовали от своих предков.
Если бы первые четвероногие жили в пресной воде, и если бы они потеряли способность производить мочевину и использовали только аммиак, им пришлось бы вырабатывать его с нуля позже. Ни один из ныне живущих здесь рыб с лучевыми плавниками не смог этого сделать, поэтому маловероятно, что четвероногие тоже поступили бы так. Наземные животные который может производить только аммиак, должен был бы постоянно пить, что сделало бы жизнь на суше невозможной (существует несколько исключений, поскольку некоторые земные мокрицы могут выделять азотистые отходы в виде газообразного аммиака). Вероятно, это также было проблемой вначале, когда четвероногие начали проводить время без воды, но в конечном итоге система мочевины стала полностью доминировать. Из-за этого маловероятно, что они появились в пресной воде (если только они сначала не мигрировали в пресноводные среды обитания, а затем мигрировали на сушу вскоре после этого, они все еще сохраняли способность производить мочевину), хотя некоторые виды никогда не покидали воду или не возвращались в нее. могли, конечно, адаптироваться к пресноводным озерам и рекам.
Легкие
Теперь ясно, что общий предок костистых рыб (Osteichthyes) имел примитивное дыхание воздухом. легкое - позже превратился в плавательный пузырь у большинства актиноптеригов (лучеплавниковых рыб). Это говорит о том, что кроссоптериги эволюционировали на теплых мелководьях, используя свое простое легкое, когда уровень кислорода в воде стал слишком низким.
Мясистые лопастные плавники, опирающиеся на кости, а не на ребристые плавники, по-видимому, были наследственной чертой всех костистых рыб (Osteichthyes ). Лопастоперые предки четвероногих развили их дальше, в то время как предки лучевых рыб (Актиноптеригии ) развили свои плавники в другом направлении. Самая примитивная группа актиноптеригов - бихиры, еще есть мясистые лобные плавники.
Окаменелости ранних четвероногих
9 роды девонских четвероногих, некоторые из них известны в основном или полностью из более низких челюсть материал. Все, кроме одного, были из Лауразийский суперконтинент, в которую вошли Европа, Северная Америка и Гренландия. Единственное исключение - одиночный Гондванец род, Метаксигнатус, который был найден в Австралия.
Первый девонский четвероногий, идентифицированный с Азия был признан из ископаемое jawbone сообщили в 2002 году. Китайский четвероногий Sinostega pani был обнаружен среди окаменелых тропических растений и рыб с плавниками в красном песчаник отложения Нинся Хуэй Автономный район северо-западного Китая. Это открытие существенно расширило географический ареал этих животных и подняло новые вопросы о мировом распространении и большом таксономическом разнообразии, которого они достигли за относительно короткое время.










Эти самые ранние четвероногие не были наземными. Самые ранние подтвержденные земные формы известны с раннего Каменноугольный отложения, примерно 20 миллионов лет спустя. Тем не менее, они могли проводить очень короткие периоды без воды и использовали бы свои ноги, чтобы прокладывать себе путь через воду. грязь.
Почему они вообще пошли на сушу, все еще обсуждается. Одной из причин могло быть то, что маленькие подростки, завершившие метаморфоза имел все необходимое, чтобы использовать то, что предлагала земля.Уже адаптированы для дыхания воздухом и передвижения по мелководью вблизи суши в качестве защиты (так же, как современные рыбы и земноводные часто проводят первую часть своей жизни в относительной безопасности мелководья, например мангровые заросли леса), две очень разные ниши частично перекрывали друг друга, между ними находилась молодая молодь. Один из них был переполнен и опасен, в то время как другой был намного безопаснее и менее многолюдным, предлагая меньшую конкуренцию за ресурсы. Наземная ниша также была гораздо более сложным местом для в первую очередь водных животных, но из-за того, как работают эволюция и давление отбора, те молодые особи, которые могли бы воспользоваться этим, будут вознаграждены. Как только они закрепятся на суше, благодаря их предварительной адаптации, благоприятные изменения в их потомках постепенно приведут к продолжению эволюции и диверсификации.
В то время множество беспозвоночных, ползающих по суше и у воды, во влажной почве и мокрой подстилке, служило источником пищи. Некоторые были даже достаточно большими, чтобы поедать маленьких четвероногих, но земля была свободна от опасностей, типичных для воды.
С воды на сушу
Первоначально совершая лишь предварительные набеги на сушу, четвероногие со временем адаптировались к земной среде и проводили более длительные периоды вдали от воды. Также возможно, что взрослые особи начали проводить некоторое время на суше (так как изменения скелета у ранних четвероногих животных, таких как Ихтиостега советует) погреться на солнышке у кромки воды[нужна цитата ], в остальном они в основном водные.
Четвероногие каменноугольные породы
До 1990-х годов в летописи окаменелостей позднедевонских четвероногих и повторного появления окаменелостей четвероногих в узнаваемом среднем возрасте наблюдался разрыв в 30 миллионов лет.Каменноугольный амфибия родословные. Это упоминалось как "Промежуток Ромера ", который в настоящее время охватывает период примерно от 360 до 345 миллионов лет назад (девонско-каменноугольный переход и ранний период Миссисипи), после палеонтолог кто это узнал.
Во время «разрыва» у четвероногих развились позвоночники, а также конечности с пальцами и другие приспособления для земной жизни. Уши, черепа и позвоночный все столбцы тоже претерпели изменения. Количество цифр на Руки и ступни стали стандартизированными до пяти, поскольку линии с большим количеством пальцев вымерли. Таким образом, те очень немногие окаменелости четвероногих, найденные в этом «промежутке», тем больше ценятся палеонтологами, потому что они документируют эти значительные изменения и разъясняют их историю.
Переход от водных рыб с лопастными плавниками к земноводным, дышащим воздухом, был значительным и фундаментальным в истории эволюции. позвоночные. Чтобы организм жил в сила тяжести -нейтральная водная среда, затем колонизация той, которая требует, чтобы организм поддерживал весь свой вес и обладал механизмом для смягчения обезвоживания, требовал значительной адаптации или экстаптации в целом план тела как по форме, так и по функциям. Эриопс, пример животного, сделавшего такую адаптацию, улучшило многие черты, присущие его предкам-рыбам. Крепкий конечности поддерживал и транспортировал свое тело, находясь вне воды. Толстее, сильнее позвоночник не давала его телу провисать под собственным весом. Кроме того, в результате изменения формы рудиментарных костей челюсти рыб начало развиваться рудиментарное среднее ухо, которое соединяется с внутреннее ухо рыб, позволяя Эриопс чтобы усилить, и поэтому лучше понять, в воздухе звук.
Посредством Визейский На стадии (середина карбона) ранние четвероногие разошлись по крайней мере на три или четыре основные ветви. Некоторые из этих различных ветвей представляют собой предков всех живых четвероногих. Это означает, что общий предок всех ныне живущих четвероногих, вероятно, жил в каменноугольном периоде. Под узким кладистический По определению Tetrapoda (также известного как Crown-Tetrapoda), которое включает в себя только потомков этого общего предка, четвероногие впервые появились в каменноугольном периоде. Узнаваемые ранние четвероногие (в широком смысле) представляют темноспондилы (например. Эриопс ) лепоспондилы (например. Диплокаулус ), антракозавры, которые были родственниками и предками Амниота, и, возможно, бафетиды, которые, как полагают, связаны с темноспондилами и статус которых в качестве основной ветви пока не решен. В зависимости от того, за какими авторитетами следует, современные амфибии (лягушки, саламандры и цецилии ), скорее всего, происходят от темноспондилов или лепоспондилов (или, возможно, от обоих, хотя сейчас это меньшинство).
Первый амниот (клада позвоночных, которая сегодня включает рептилии, млекопитающие, и птицы ) известны с самого начала Поздний карбон. Посредством Триасовый, эта группа уже излучалась в самые ранние млекопитающие, черепахи, и крокодилы (ящерицы и птицы появился в Юрский, и змеи в Меловой ). Это резко контрастирует с (возможно, четвертым) Каменноугольный группа, бафетиды, которые не оставили никаких сохранившихся линий.
Обрушение тропических лесов каменноугольного периода
Амфибии и рептилии сильно пострадали от Обрушение тропических лесов каменноугольного периода (CRC), событие вымирания, которое произошло ~ 307 миллионов лет назад. Каменноугольный период издавна ассоциировался с густыми парными болотами и влажными тропическими лесами.[58] Поскольку растения составляют основу почти всех экосистем Земли, любые изменения в их распространении всегда в той или иной степени влияли на жизнь животных. Внезапный коллапс жизненно важной экосистемы тропических лесов глубоко повлиял на разнообразие и численность основных групп четвероногих, которые полагались на нее.[59] CRC, который был частью одного из двух самых разрушительных исчезновений растений в истории Земли, был самоусиливающимся и очень быстрым изменением окружающей среды, в результате которого климат во всем мире в целом стал намного суше и прохладнее (хотя в настоящее время проводится много новой работы. чтобы лучше понять мелкозернистые исторические изменения климата в переходный период между каменноугольным и пермским периодом и их возникновение.[60]).
Последовавшее за этим сокращение растений во всем мире в результате трудностей, с которыми растения столкнулись при адаптации к новому климату, вызвало прогрессирующую фрагментацию и коллапс экосистем тропических лесов. Это усилило и, таким образом, еще больше ускорило коллапс, резко сократив количество животных, которые могли поддерживаться сокращающимися экосистемами в то время. Результатом этого сокращения количества животных стало падение глобального уровня углекислого газа, что еще больше повлияло на растения.[61] Засушливость и падение температуры, возникшие в результате этого безудержного сокращения растений и уменьшения количества первичных парниковых газов, привели к тому, что Земля быстро вступила в серию интенсивных ледниковых периодов.[58]
Это сказалось, в частности, на амфибиях по разным причинам. Огромное падение уровня моря из-за того, что большее количество мировой воды было заперто в ледниках, сильно повлияло на распределение и размер полуводных экосистем, которым благоприятствовали земноводные, а значительное похолодание климата еще больше сузило количество новых территорий, благоприятных для земноводных. Учитывая, что среди отличительных черт земноводных - обязательное возвращение в водоем для откладывания яиц, нежная кожа, склонная к высыхание (тем самым часто требуя от земноводных находиться относительно близко к воде на протяжении всей своей жизни) и репутации одного из видов-лидеров нарушенных экосистем из-за низкой устойчивости к экологическим изменениям,[62] земноводные были особенно опустошены, а Лабиринтодонты среди групп дела обстоят хуже всего. Напротив, рептилии, чьи амниотический Яйца имеют мембрану, которая обеспечивает газообмен вне воды и, таким образом, их можно откладывать на суше - они лучше приспособились к новым условиям. Рептилии быстрее вторгались в новые ниши и начали диверсифицировать свой рацион, превратившись в травоядных и плотоядных, вместо того, чтобы питаться исключительно насекомыми и рыбой.[63] Между тем, сильно пострадавшие амфибии просто не могли опередить рептилий в освоении новых экологических ниш.[64] и поэтому они были вынуждены передать факел эволюции четвероногих все более успешным и быстро распространяющимся рептилиям.
Пермские четвероногие
в Пермский период период: включены ранние клады амфибий (лабиринтодонтов) темноспондил и антракозавр; пока амниот клады включали Sauropsida и Synapsida. Со временем зауропсида превратилась в сегодняшнюю рептилии и птицы; тогда как Synapsida превратится в сегодняшнюю млекопитающие. Вовремя Пермский период однако различие было менее четким - фауну амниот обычно описывали как рептилия или как млекопитающее рептилии. Последние (синапсиды) были наиболее важными и успешными пермскими животными.
В конце перми произошел крупный оборот фауны во время Пермско-триасовое вымирание: наверное самый тяжелый событие массового вымирания из фанерозойский. Произошла длительная потеря видов из-за многократных импульсов вымирания.[65] Многие из когда-то больших и разнообразных групп вымерли или сильно сократились.
Мезозойские четвероногие
Жизнь на Земле, казалось, быстро восстановилась после пермского вымирания, хотя в основном это происходило в форме таксоны бедствий такие как выносливые Lystrosaurus. Специализированным животным, которые сформировали сложные экосистемы с высоким биоразнообразием, сложными пищевыми сетями и множеством ниш, требовалось гораздо больше времени для восстановления.[65] Текущие исследования показывают, что это долгое восстановление было связано с последовательными волнами вымирания, которые препятствовали восстановлению, а также с длительным экологическим стрессом для организмов, который продолжался в раннем триасе. Недавние исследования показывают, что восстановление началось только в начале среднего триаса, через 4-6 млн лет после исчезновения;[66] и некоторые авторы считают, что восстановление было завершено только через 30 миллионов лет после вымирания P-Tr, то есть в конце триаса.[65]
Небольшая группа рептилий, диапсиды, начали диверсифицироваться в триасовом периоде, особенно динозавры. К концу мезозоя большая лабиринтодонт группы, которые впервые появились в палеозое, такие как темноспондилы и рептилоидные амфибии вымерли. Все нынешние основные группы зауропсид эволюционировали в мезозое, с птицы впервые появившись в Юрский как производный от теропод динозавры. Многие группы синапсиды Такие как аномодонты и тероцефалы которые когда-то составляли доминирующую наземную фауну перми, также вымерли в мезозое; однако в триасе одна группа (Цинодонтия ) дал начало таксону-потомку Млекопитающие, которые пережили мезозой, чтобы позже разнообразить в течение кайнозоя.
Кайнозойские четвероногие
В Кайнозойский эра началась с концом Мезозойский эпоха и Меловой эпоха; и продолжается по сей день. Начало кайнозоя ознаменовалось началом Мелово-палеогеновое вымирание во время которого все нептичьи динозавры вымерли. Кайнозой иногда называют «эпохой Млекопитающие ".
В мезозое прототипом млекопитающего был маленький ночной образ жизни насекомоядное животное что-то вроде землеройка. Из-за своих ночных привычек большинство млекопитающих потеряли цветовое зрение, и значительно улучшили их чувство слушание. Это происхождение сформировало всех современных млекопитающих. Приматы позже заново развил цветовое зрение.
Вовремя Палеоцен и эоцен, большинство млекопитающих остались мелкими (до 20 кг). Прохладный климат в Олигоцен и Миоцен, а расширение луга способствовал развитию более крупных видов млекопитающих.
Ratites бежать, и пингвины плавать и гулять: но большинство птицы довольно маленькие и могут летать. Некоторые птицы используют свою способность летать, чтобы совершить эпическое пересечение земного шара. миграции, а другие, такие как фрегат птицы месяцами летают над океанами.
Летучие мыши также улетели, и вместе с китообразные разработали эхолокация или же сонар.
Киты, уплотнения, ламантины, и морские выдры вернулись в океан и водный Стиль жизни.
Обширные стада жвачный копытные заселить луга и леса. Хищники эволюционировали, чтобы держать под контролем популяции стадных животных.
Сохранившиеся (живые) четвероногие
После большого обновления фауны в конце мезозоя осталось только шесть основных групп четвероногих, каждая из которых также включает многих вымерший группы:
- Лиссамфибия: лягушки и жабы, тритоны и саламандры, и цецилии
- Testudines: черепахи и черепахи
- Лепидозаврия: Туатары, ящерицы, амфисбайцы и змеи
- Крокодилы: крокодилы, аллигаторы, кайманы и гавиалы
- Neornithes: современные птицы
- Млекопитающие: млекопитающие
Рекомендации
- ^ а б Шубин, Н. (2008). Ваша внутренняя рыба: путешествие в 3,5 миллиарда лет истории человеческого тела. Нью-Йорк: Книги Пантеона. ISBN 978-0-375-42447-2.
- ^ Клак, Дженнифер А. (1997). «Следы девонских четвероногих и создатели путей; обзор окаменелостей и следов». Палеогеография, палеоклиматология, палеоэкология. 130 (1–4): 227–250. Bibcode:1997ППП ... 130..227С. Дои:10.1016 / S0031-0182 (96) 00142-3.
- ^ Лаурин, М. (2010). Как позвоночные животные вышли из воды. Беркли, Калифорния, США: Калифорнийский университет Press. ISBN 978-0-520-26647-6.
- ^ Кановилл, Аврора; Лорен, Мишель (2010). «Эволюция микроанатомии плечевой кости и образа жизни у амниот, и некоторые комментарии к палеобиологическим выводам». Биологический журнал Линнеевского общества. 100 (2): 384–406. Дои:10.1111 / j.1095-8312.2010.01431.x.
- ^ Лорен, Мишель; Кановилл, Аврора; Quilhac, Александра (2009). «Использование палеонтологических и молекулярных данных в супердеревьях для сравнительных исследований: на примере лиссамфибийской бедренной микроанатомии». Журнал анатомии. 215 (2): 110–123. Дои:10.1111 / j.1469-7580.2009.01104.x. ЧВК 2740958. PMID 19508493.
- ^ а б Лонг Дж. А., Гордон М. С. (2004). «Величайший шаг в истории позвоночных: палеобиологический обзор перехода рыба-четвероногие». Physiol. Biochem. Zool. 77 (5): 700–19. Дои:10.1086/425183. PMID 15547790. S2CID 1260442. В архиве из оригинала на 2016-04-12. Получено 2014-03-09. как PDF В архиве 2013-10-29 в Wayback Machine
- ^ Уэллс, Х. (1922). «Глава IV: Эпоха рыб». Краткая история мира. Макмиллан. ISBN 978-1-58734-075-8. Архивировано из оригинал на 2014-02-01. Получено 2014-03-09..
- ^ Колберт, Эдвин Х. (1969). Эволюция позвоночных (2-е изд.). Джон Вили и сыновья. стр.49–53.
- ^ Бентон 2005, п. 67
- ^ «Эволюция позвоночных началась в лагунах». В архиве из оригинала на 2018-11-12. Получено 2018-11-12.
- ^ «Прибрежная колыбель раннего разнообразия позвоночных | Наука». В архиве из оригинала на 2019-03-08. Получено 2018-11-12.
- ^ Веколи, Марко; Клеман, Гаэль; Мейер-Берто, Б. (2010). Процесс террестриализации: моделирование сложных взаимодействий на границе биосферы и геосферы. ISBN 9781862393097. В архиве из оригинала на 2018-11-12. Получено 2018-11-12.
- ^ «Эвригалинная экология ранних четвероногих по стабильным изотопам - Природа». В архиве из оригинала от 23.03.2019. Получено 2018-11-12.
- ^ Стин, Йохан Б. (1970). «Плавательный пузырь как гидростатический орган». Физиология рыб. 4. Сан-Диего, Калифорния: Academic Press, Inc., стр. 413–443. ISBN 9780080585246. В архиве из оригинала от 02.03.2016. Получено 2016-01-27.
- ^ Виделер, Дж. Дж. (1993). Рыба плавание. Нью-Йорк: Чепмен и Холл.
- ^ а б Даль Т.В., Хаммарлунд ЕС, Анбар А.Д. и др. (Октябрь 2010 г.). «Девонский подъем атмосферного кислорода связан с излучением наземных растений и крупных хищных рыб». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 107 (42): 17911–5. Bibcode:2010PNAS..10717911D. Дои:10.1073 / pnas.1011287107. ЧВК 2964239. PMID 20884852.
- ^ Вакер-Суньер Р., Дуарте К.М. (октябрь 2008 г.). «Пороги гипоксии для морского биоразнообразия». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 105 (40): 15452–7. Bibcode:2008ПНАС..10515452В. Дои:10.1073 / pnas.0803833105. ЧВК 2556360. PMID 18824689.
- ^ Gray, J .; Wu, R .; Или, Ю. (2002). Влияние гипоксии и обогащения органическими веществами на прибрежную морскую среду. Серия «Прогресс морской экологии». 238. С. 249–279. Bibcode:2002MEPS..238..249G. Дои:10,3354 / meps238249.
- ^ Армбрустер, Джонатан В. (1998). «Модификации пищеварительного тракта для удержания воздуха у лорикариевых и сколоплацидных сомов» (PDF). Копея. 1998 (3): 663–675. Дои:10.2307/1447796. JSTOR 1447796. В архиве (PDF) из оригинала 26.03.2009. Получено 25 июн 2009.
- ^ Лонг, Дж. (1990). «Гетерохрония и происхождение четвероногих». Lethaia. 23 (2): 157–166. Дои:10.1111 / j.1502-3931.1990.tb01357.x.
- ^ Ромер, А. (1949). Тело позвоночного. Филадельфия: W.B. Сондерс. (2-е изд. 1955; 3-е изд. 1962; 4-е изд. 1970)
- ^ а б Kent, G.C .; Миллер, Л. (1997). Сравнительная анатомия позвоночных (8-е изд.). Дубюк: Wm. C. Brown Publishers. ISBN 978-0-697-24378-2.
- ^ Hildebran, M .; Гослоу, Г. (2001). Анализ структуры позвоночных (5-е изд.). Нью-Йорк: Джон Вили. ISBN 978-0-471-29505-1.
- ^ а б Фернандес, Мариса Нарцисо; да Круз, Андре Луис; да Кошта, Оскар Тадеу Феррейра; Перри, Стивен Франклин (сентябрь 2012 г.). «Морфометрическое разделение площади дыхательной поверхности и диффузионной способности жабр и плавательного пузыря у молодых амазонских дышащих воздухом рыб, Arapaima gigas». Микрон (Оксфорд, Англия: 1993). 43 (9): 961–970. Дои:10.1016 / j.micron.2012.03.018. ISSN 1878-4291. PMID 22512942.
- ^ Brauner, C.J .; Matey, V .; Wilson, J.M .; Bernier, N.J .; Вал, А. Л. (2004-04-01). «Изменение функций органов в ходе эволюции дыхания воздухом; выводы из Arapaima gigas, облигатной костистости с Амазонки, дышащей воздухом». Журнал экспериментальной биологии. 207 (9): 1433–1438. Дои:10.1242 / jeb.00887. ISSN 0022-0949. PMID 15037637.
- ^ Дэниелс, Кристофер Б .; Оргейг, Сандра; Салливан, Люси С .; Линг, Николас; Беннетт, Майкл Б.; Шюрх, Самуэль; Вал, Адальберто Луис; Браунер, Колин Дж. (Сентябрь 2004 г.). «Происхождение и эволюция системы поверхностно-активных веществ у рыб: понимание эволюции легких и плавательного пузыря». Физиологическая и биохимическая зоология. 77 (5): 732–749. CiteSeerX 10.1.1.385.9019. Дои:10.1086/422058. ISSN 1522-2152. PMID 15547792. S2CID 9889616.
- ^ Оргейг, Сандра; Моррисон, Жанна Л .; Дэниелс, Кристофер Б. (31 августа 2011 г.). «Пренатальное развитие сурфактантной системы легких и влияние гипоксии». Респираторная физиология и нейробиология. 178 (1): 129–145. Дои:10.1016 / j.resp.2011.05.015. ISSN 1878-1519. PMID 21642020. S2CID 41126494.
- ^ Hsia, Connie C.W .; Шмитц, Анке; Ламбертц, Маркус; Перри, Стивен Ф .; Майна, Джон Н. (апрель 2013 г.). «Эволюция дыхания воздухом: гомеостаз кислорода и переходы от воды к земле и небу». Комплексная физиология. 3 (2): 849–915. Дои:10.1002 / cphy.c120003. ISSN 2040-4603. ЧВК 3926130. PMID 23720333.
- ^ Хоффман, М .; Taylor, B.E .; Харрис, М. Б. (апрель 2016 г.). «Эволюция легочного дыхания у примитивного позвоночного без легких». Респираторная физиология и нейробиология. 224: 11–16. Дои:10.1016 / j.resp.2015.09.016. ISSN 1878-1519. ЧВК 5138057. PMID 26476056.
- ^ Панчен, А. Л. (1967). «Ноздри хоанатных рыб и ранних четвероногих». Биол. Rev. 42 (3): 374–419. Дои:10.1111 / j.1469-185X.1967.tb01478.x. PMID 4864366. S2CID 36443636.
- ^ Чжу, Мин; Альберг, Пер Э. (2004). «Происхождение внутренней ноздри четвероногих». Природа. 432 (7013): 94–7. Bibcode:2004Натура 432 ... 94Z. Дои:10.1038 / природа02843. PMID 15525987. S2CID 4422813. Архивировано из оригинала
| архив-url =требует| url =(помощь) 20 апреля 2009 г. Сложить резюме – инновации-отчет (4 ноября 2004 г.).| дата доступа =требует| url =(помощь) - ^ Коутс, Майкл I .; Джеффри, Джонатан Э .; Рута, Марселла (2002). «От плавников до конечностей: что говорят окаменелости» (PDF). Эволюция и развитие. 4 (5): 390–401. Дои:10.1046 / j.1525-142X.2002.02026.x. PMID 12356269. S2CID 7746239. Архивировано из оригинал (PDF) на 2010-06-10. Получено 18 февраля, 2013.
- ^ Геологическая служба Канады (2008-02-07). «Прошлые жизни: Хроники канадской палеонтологии: Евстеноптерон - принц Мигуаша». Архивировано из оригинал на 2004-12-11. Получено 2009-02-10.
- ^ Meunier, François J .; Лорен, Мишель (Январь 2012 г.). "Микроанатомическое и гистологическое исследование длинных костей плавников девонского саркоптерига" Eusthenopteron foordi". Acta Zoologica. 93 (1): 88–97. Дои:10.1111 / j.1463-6395.2010.00489.x.
- ^ Ahlberg, P.E .; Йохансон, З. (1998). «Остеолепиформ и предки четвероногих» (PDF). Природа. 395 (6704): 792–794. Bibcode:1998Натура.395..792А. Дои:10.1038/27421. S2CID 4430783. Архивировано из оригинал (PDF) в 2014-11-24. Получено 2014-03-09.
- ^ Мой-Томас, Дж. А. (1971). Палеозойские рыбы (2-е изд., Обширно перераб. Ред.). Филадельфия: Сондерс. ISBN 978-0-7216-6573-3.
- ^ Эндрюс, С. М. (январь 1985 г.). «Ризодонтовая кроссоптериская рыба из Динантиана в Фулдене, Берикшир, Шотландия, с переоценкой этой группы». Труды Королевского общества Эдинбурга: науки о Земле. 76 (1): 67–95. Дои:10.1017 / S0263593300010324.
- ^ Рута, Марчелло; Джеффри, Джонатан Э .; Коутс, Майкл И. (2003). «Супердерево ранних четвероногих». Труды Королевского общества B. 270 (1532): 2507–16. Дои:10.1098 / rspb.2003.2524. ЧВК 1691537. PMID 14667343.
- ^ Университет Монаша. "Окаменелая находка в Западной Австралии переписывает эволюцию наземных млекопитающих В архиве 2017-08-21 в Wayback Machine. "ScienceDaily 19 октября 2006 г. По состоянию на 11 марта 2009 г.
- ^ «Тетрапода». Палеос интернет сайт. Архивировано из оригинал на 2013-03-29. Получено 11 октября 2012.
Еще более близкое родство было Пандерихтис, у которого даже была хоана. Эти рыбы использовали свои плавники как весла в мелководных местах обитания, забитых растениями и детритом.
- ^ «Окаменелости рыб возрастом 375 миллионов лет проливают свет на эволюцию от плавников до конечностей». 2014-01-14. В архиве из оригинала от 07.04.2014. Получено 2014-05-31.
- ^ Эшли-Росс, М. А .; Hsieh, S.T .; Gibb, A.C .; Блоб, Р. В. (2013). «Вторжения на сушу позвоночных - прошлое, настоящее и будущее: введение в симпозиум». Интегративная и сравнительная биология. 53 (2): 192–196. Дои:10.1093 / icb / ict048. PMID 23660589. В архиве с оригинала от 13.10.2014. Получено 2014-05-31.
- ^ Шнайдер, Игорь; Шубин, Нил Х. (декабрь 2012 г.). "Изготовление конечностей из ласт". Клетка развития. 23 (6): 1121–1122. Дои:10.1016 / j.devcel.2012.11.011. PMID 23237946.
- ^ Zhang, J .; Wagh, P .; Guay, D .; Sanchez-Pulido, L .; Padhi, B.K .; Корж, В .; Андраде-Наварро, М. А .; Акименко М.А. (2010). «Потеря белков актинотрихии рыб и переход от плавников к конечностям». Природа. 466 (7303): 234–237. Bibcode:2010Натура.466..234Z. Дои:10.1038 / природа09137. PMID 20574421. S2CID 205221027.
- ^ Ijspeert, A.J .; Crespi, A .; Ryczko, D .; Кабельгуен, Ж.-М. (9 марта 2007 г.). «От плавания к ходьбе с роботом-саламандрой, управляемым моделью спинного мозга». Наука. 315 (5817): 1416–1420. Bibcode:2007Научный ... 315.1416I. Дои:10.1126 / science.1138353. PMID 17347441. S2CID 3193002. В архиве из оригинала 16 января 2020 г.. Получено 7 декабря 2019.
- ^ Standen, Emily M .; Du, Trina Y .; Ларссон, Ханс К. Э. (27 августа 2014 г.). «Пластичность развития и происхождение четвероногих». Природа. 513 (7516): 54–58. Bibcode:2014Натура.513 ... 54S. Дои:10.1038 / природа13708. PMID 25162530. S2CID 1846308.
- ^ Стефани Э. Пирс; Дженнифер А. Клак; Джон Р. Хатчинсон (2012). «Трехмерная подвижность суставов конечностей у ранних четвероногих. Ихтиостега". Природа. 486 (7404): 524–527. Bibcode:2012Натура 486..523П. Дои:10.1038 / природа11124. PMID 22722854. S2CID 3127857.
- ^ Clack 2002, стр. 86–7
- ^ а б Гжегож Недзведски; Петр Шрек; Катажина Наркевич; Марек Наркевич; Пер Э. Альберг (2010). «Пути тетрапод раннего среднего девона в Польше». Природа. 463 (7277): 43–8. Bibcode:2010Натура.463 ... 43N. Дои:10.1038 / природа08623. PMID 20054388. S2CID 4428903.
- ^ Рекс Далтон (6 января 2010 г.). «Открытие отодвигает дату появления первого четвероногого животного». Новости природы. В архиве из оригинала 14.01.2010. Получено 8 января, 2010.
- ^ Clack 2012, п. 140
- ^ «Маленький шаг для двоякодышки, большой шаг в эволюции ходьбы». В архиве из оригинала на 2017-07-03. Получено 2018-02-28.
- ^ King, H.M .; Шубин, Н. Х .; Coates, M. I .; Хейл, М. Э. (2011). «Поведенческие свидетельства эволюции ходьбы и прыжков до появления на суше у саркоптерийских рыб». Труды Национальной академии наук. 108 (52): 21146–21151. Bibcode:2011PNAS..10821146K. Дои:10.1073 / pnas.1118669109. ЧВК 3248479. PMID 22160688. В архиве из оригинала 30.05.2014. Получено 2014-03-09.
- ^ Реталлак, Грегори (май 2011 г.). "Гипотеза Вудленда для эволюции девонских тетрапод" (PDF). Журнал геологии. Издательство Чикагского университета. 119 (3): 235–258. Bibcode:2011JG .... 119..235R. Дои:10.1086/659144. S2CID 128827936. В архиве (PDF) из оригинала 17.05.2013. Получено 1 января, 2012.
- ^ «Появляется новая теория о том, где некоторые рыбы превратились в четвероногих существ». Наука. 28 декабря 2011 г. Архивировано с оригинал на 2016-03-04. Получено 17 января, 2013.
- ^ Джордж р. Макги-младший (12 ноября 2013 г.). Когда вторжение на землю провалилось: наследие девонских вымираний. ISBN 9780231160575. В архиве с оригинала на 2019-12-27. Получено 2016-03-01.
- ^ «Исследовательский проект: Среднепалеозойский биотический кризис: определение траектории эволюции тетрапод». В архиве из оригинала 12.12.2013. Получено 2014-05-31.
- ^ а б Dimichele, William A .; Сесил, К. Блейн; Montañez, Isabel P .; Фалькон-Лэнг, Ховард Дж. (2010). «Циклические изменения в палеоклимате Пенсильвании и влияние на флористическую динамику в тропической Пангеи». Международный журнал угольной геологии. 83 (2–3): 329–344. Дои:10.1016 / j.coal.2010.01.007.
- ^ Дэвис, Нил С .; Гиблинг, Мартин Р. (2013). «Отложения рек каменноугольного периода: продолжающееся влияние эволюции наземных растений на аллювиальные процессы и палеозойские экосистемы». Обзоры наук о Земле. 120: 40–79. Bibcode:2013ESRv..120 ... 40D. Дои:10.1016 / j.earscirev.2013.02.004.
- ^ Табор, Нил Дж .; Поульсен, Кристофер Дж. (2008). «Палеоклимат в тропических палеоширотах позднего пенсильвана – ранней перми: обзор климатических индикаторов, их распределения и связи с палеофизиографическими климатическими факторами». Палеогеография, палеоклиматология, палеоэкология. 268 (3–4): 293–310. Bibcode:2008ППП ... 268..293Т. Дои:10.1016 / j.palaeo.2008.03.052.
- ^ Гиблинг, М.Р .; Davies, N.S .; Falcon-Lang, H.J .; Bashforth, A.R .; Dimichele, W.A .; Rygel, M.C .; Иелпи, А. (2014). «Палеозойская совместная эволюция рек и растительности: синтез современных знаний». Труды ассоциации геологов. 125 (5–6): 524–533. Дои:10.1016 / j.pgeola.2013.12.003.
- ^ Purves, Уильям К .; Orians, Gordon H .; Хеллер, Х. Крейг (1995). Жизнь, наука о биологии (4-е изд.). Сандерленд, Массачусетс, США: Sinauer Associates. стр.622 –625. ISBN 978-0-7167-2629-6.
- ^ Sahney, S .; Benton, M.J .; Фалькон-Лэнг, Х.Дж. (2010). «Коллапс тропических лесов вызвал диверсификацию пенсильванских четвероногих в Европе». Геология. 38 (12): 1079–1082. Bibcode:2010Гео .... 38.1079S. Дои:10.1130 / G31182.1.
- ^ Пирсон, Марианна Р .; Бенсон, Роджер Б.Дж .; Апчерч, Пол; Fröbisch, Jörg; Каммерер, Кристиан Ф. (2013). «Реконструкция разнообразия ранних наземных травоядных четвероногих». Палеогеография, палеоклиматология, палеоэкология. 372: 42–49. Bibcode:2013ГПП ... 372 ... 42П. Дои:10.1016 / j.palaeo.2012.11.008.
- ^ а б c Sahney, S .; Бентон, М.Дж. (2008). «Восстановление после самого глубокого массового вымирания всех времен» (PDF). Труды Королевского общества B: биологические науки. 275 (1636): 759–65. Дои:10.1098 / rspb.2007.1370. ЧВК 2596898. PMID 18198148. В архиве (PDF) из оригинала от 22.02.2011. Получено 2014-03-09.
- ^ Lehrmann, D.J .; Ramezan, J .; Bowring, S.A .; и другие. (Декабрь 2006 г.). «Время восстановления от конца пермского вымирания: геохронологические и биостратиграфические ограничения из южного Китая». Геология. 34 (12): 1053–6. Bibcode:2006Geo .... 34.1053L. Дои:10.1130 / G22827A.1.
внешняя ссылка
 СМИ, связанные с Эволюция четвероногих в Wikimedia Commons
СМИ, связанные с Эволюция четвероногих в Wikimedia Commons

{kind=link}