Австралопитек седиба - Australopithecus sediba
| Австралопитек седиба | |
|---|---|
 | |
| Реконструированный скелет MH1 на Музей естественной истории, Лондон | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Haplorhini |
| Инфразаказ: | Симиобразные |
| Семья: | Гоминиды |
| Подсемейство: | Гоминины |
| Племя: | Гоминини |
| Род: | Австралопитек |
| Разновидность: | А. седиба |
| Биномиальное имя | |
| Австралопитек седиба | |
Австралопитек седиба является вымерший разновидность из австралопитеки выздоровел от Пещера Малапа, Колыбель человечества, Южная Африка. Он известен по частичному скелету молоди, голотип MH1, и частичный скелет взрослой женщины, паратип MH2. Они датируются примерно 1,98 миллиона лет назад в Ранний плейстоцен и сосуществовали с Paranthropus robustus и Homo ergaster /H. erectus. Малапа интерпретируется как естественная смертельная ловушка, основание длинной вертикальной шахты, в которую существа могли случайно упасть. А. седиба изначально был описан как потенциальный предок человека и, возможно, прародитель Гомо, но это оспаривается, и это может также представлять поздно выжившую популяцию или родственные виды из A. africanus ранее населявшие этот район.
MH1 имеет объем мозга около 420–440 куб. См, как и у других австралопитеков. Лицо MH1 поразительно похоже на Гомо вместо других австралопитеков, с менее выраженным надбровным дугом, скулами и прогнатизм (размер лица выступает), и есть свидетельства небольшого подбородка. Однако такие характеристики могут быть связаны с молодостью и утрачены со зрелостью. Зубы у австралопитека довольно маленькие. MH1 оценивается в 130 см (4 фута 3 дюйма) в высоту, что соответствует росту взрослого человека в 150–156 см (4 фута 11 дюймов – 5 футов 1 дюйм). По оценкам, MH1 и MH2 имели примерно одинаковый вес - 30–36 кг (66–79 фунтов). Как и другие австралопитеки, А. седиба считается, что у него была узкая и похожая на обезьяна верхняя часть груди, но широкая и похожая на человеческую нижнюю часть. Как и у других австралопитеков, анатомия руки предполагает определенную степень лазания и древесный поведение. В таз указывает А. седиба был способен к походке, подобной человеческой, но ступня указывает на особую походку, не продемонстрированную ни у одного другого гоминина, включающую гиперпронирование лодыжки и, как следствие, вращение ноги внутрь при отталкивании. Этот набор приспособлений может представлять собой компромисс между привычными двуногие и древесину.
А. седиба кажется, ел только C3 лесные растения, такие как травы и осока, плоды, листья и кора. Это сильно отличается от других ранних гоминины которые съели смесь C3 и обильный C4 растения саванны, но похожи на современную саванну шимпанзе. Ни один другой гоминин не имеет доказательств поедания коры. Такой универсал диета, возможно, позволила ему занять меньший домашний ареал, чем шимпанзе саванны. Район Малапы, возможно, был более прохладным и влажным, чем сегодня, с закрытыми лесами, окруженными более открытыми лугами.
История исследований
Образцы


Первая ископаемая находка была правильной ключица, MH1 (UW88-1), дюйм Пещера Малапа, Колыбель человечества, Южная Африка, обнаружен 9-летним Мэтью Бергером 15 августа 2008 года во время исследования раскопок, возглавляемых его отцом, южноафриканским палеоантропологом. Ли Роджерс Бергер. Дальнейшие раскопки позволили получить частичный скелет MH1, дополнительно включающий частичные фрагменты черепа и челюстной кости, а также части рук, пальцев, плеч и других частей тела. грудная клетка, позвоночник, таз, ноги и ступни. MH1 интерпретируется как молодой самец из-за явно выраженного развития надбровных дуг и собачий корни, выворот угол нижней челюсти и большие рубцы на костях.[1] Однако антропологи Уильям Кимбел и Йоэль Рак утверждают, что это ненадежные методы определения пола, и предполагают, что MH1 является женским, на основании отсутствия передних столбов (столбцов, идущих вниз вдоль носового отверстия до области вокруг рта) и слегка выпуклой подназальной части. пластина, используя методы определения пола для A. africanus.[2] MH1 получил прозвище «Карабо», что в переводе с английского означает «ответ». Тсвана, 17-летней Омфеметсе Кипиле из Школа Святой Марии, Йоханнесбург, в конкурсе имен. Она выбрала это название, потому что «окаменелость представляет собой решение для понимания происхождения человечества».[3]
Другой частичный скелет, взрослый MH2, был обнаружен Ли 4 сентября 2008 года с изолированными верхними зубами, частичной челюстной костью, почти полной правой рукой, правой рукой. лопатка, а также фрагменты плеч, правой руки, позвоночника, ребер, таза, коленный сустав, и ноги. В лобковая кость широкая и квадратная, а рубцы на мышцах тела от слабых до умеренных, что позволяет предположить, что MH2 принадлежит женщине.[1]


Присутствие видов, которые эволюционировали после 2,36 миллиона лет назад и вымерли около 1,5 миллиона лет назад, указывает на то, что А. седиба слой датируется временем в пределах этого интервала во время Ранний плейстоцен. Уран-свинец датирование из Flowstone покрывающий слой дал дату 2,026 ± 0,021 миллиона лет назад. С помощью археомагнитное датирование, отложения имеют нормальный магнитная полярность (в отличие от обратной магнитной полярности в наши дни), и единственное время, когда это произошло в течение этого интервала, было между 1,95 и 1,78 миллиона лет назад.[4] В 2011 году датировка водоплавающего камня была более точной - 1,977 ± 0,002 миллиона лет назад, опять же с использованием уран-свинцового датирования.[5]
Тафономия


Сеть пещер вокруг Малапы состоит из длинных соединенных между собой пещерных входов в пределах 500 м × 100 м (1640 футов × 330 футов). Участок Малапа мог быть у основания системы пещер глубиной не более 30 метров (98 футов). Пещера находится на пересечении северо-северо-востока и северо-северо-запада. черт -заполненная трещина, и останки гомининов были обнаружены в разрезе 3,3 м × 4,4 м × 3,5 м (11 футов × 14 футов × 11 футов) на северо-северо-западной трещине. Слой экспонировался известняк горное дело в начале 20 века. Пещера состоит из пяти осадочный фации A – E гидроизоляции песчаник, с А. седиба извлекаются из фации D, и больше остатков гомининов из фации E. MH1 и MH2 разделены по вертикали максимум на 40 см (16 дюймов). Фация D представляет собой слой светлого цвета толщиной 1,5 метра (4,9 фута), лежащий над водопадом. Маленький пелоиды являются обычными, но сливаются в большие и неправильные группы, что указывает на то, что они отложились в условиях заболоченного грунта. Пелоиды могут представлять фекалии или почвенные микробы. Состояние сохранности MH1 и MH2 указывает на то, что они откладывались быстро, перемещались очень мало и были цементированный вскоре после депонирования в фреатический окружающая среда (в подземном потоке). Нет никаких свидетельств выгула мусора, что указывает на то, что этот район был недоступен для хищников.[4]
Все это могло указывать на то, что пещера Малапа была смертельной ловушкой с неприметными пещерными отверстиями на поверхности. Животные могли быть соблазнены запахом воды, исходящей из шахты, а плотоядных - запахом мертвых животных, и затем они упали насмерть. Большой поток обломков привел к тому, что останки осели глубже в пещере вдоль подземного ручья, возможно, из-за сильного ливня. В конце концов камера рухнула и заполнилась грязью.[4]
Классификация

В 2010 году Ли и его коллеги официально описанный виды Австралопитек седиба с MH1 в качестве голотип и MH2 паратип. Название вида "седиба"означает" фонтан "или" источник "на местном языке Язык сесото.[1] Потому что А. седиба имел много общих черт с Homo ergaster /H. erectus, особенно в области таза и ног, авторы постулировали, что А. седиба был переходное ископаемое между Австралопитек и Гомо.[1] Стоматологические особенности также наводят на мысль о некоторой близкой связи между А. седиба и предок Гомо.[6] Однако экземпляры были найдены в стратиграфическая единица датируется 1,95–1,78 миллиона лет назад, тогда как самые ранние Гомо окаменелости того времени, датированные 2,33 миллиона лет назад (H. habilis из Адар, Эфиопия).[1] В настоящее время самый старый Гомо образец LD 350-1 датируется 2,8–2,75 миллиона лет назад Леди-Герару, Эфиопия.[7] Чтобы примирить расхождение в датировке, описатели также выдвинули гипотезу, что А. седиба произошел от населения A. africanus (которые населяли тот же регион) некоторое время до гомининов Малапы, и что Гомо отделиться от А. седиба когда-нибудь после этого.[1] Это будет означать 800000 лет призрачное происхождение между A. africanus и гоминины Малапы.[2] Было также предложено, чтобы А. седиба, вместо H. habilis или же Х. рудольфенсис, был прямым предком Х. эргастер/H. erectus (самый ранний неоспоримый представитель рода Гомо), в первую очередь потому, что гоминины Малапа были датированы 1,98 млн лет назад в 2011 году, что в то время предшествовало самому раннему представителю Х. эргастер/H. erectus.[5] А. седиба теперь считается, что он был современником Х. эргастер/H. erectus и Paranthropus robustus в колыбели человечества.[8]
В качестве альтернативы, А. седиба может также представлять позднее сохранившийся морф или родственные виды из A. africanus не связанный с Гомо, что означало бы Гомо-подобные черты независимо развились в А. седиба и Гомо (гомоплазия ).[2][9][10][11][12] Летопись окаменелостей ранних Гомо малоизвестен и основан в основном на фрагментарных останках, что делает убедительные анатомические сравнения трудными, а иногда и невозможными.[12] A. africanus, A. afarensis, и А. гархи также были предложены как истинные предки Гомо, и этот вопрос очень обсуждается.[7] Кроме того, голотип - это несовершеннолетний, которого Кимбел и Рак цитируют, утверждая, что некоторые из Гомо-подобные черты лица могли быть потеряны с возрастом.[2]
Настоящая классификация австралопитеки в беспорядке. Австралопитек можно считать таксон ранга члены которых объединены сходной физиологией, а не близкими отношениями друг с другом по другим родам гомининов, и, по большей части, в значительной степени неясно, как один вид относится к другим.[13]
 |
Анатомия
Череп

Только свод черепа сохранилась MH1 объемом 363 куб. Размер самой задней части мозга оценивается в 7–10 куб. Чтобы оценить мозжечок, австралопитеки KNM-ER 23000 (Paranthropus boisei ) и Sts 19 (A. africanus) объемом 40–50 см3, а также KNM-ER 1813 (H. habilis), KNM-ER 1805 (H. habilis) и KNM-ER 1470 (Х. рудольфенсис) с объемами 55–75 см3 были использованы для оценки объема мозжечка MH1 примерно в 50 см3. Учитывая все это, MH1 мог иметь объем мозга примерно 420–440 куб. См. Это типично для австралопитеков.[1] Используя тенденции, наблюдаемые у современных приматов между размером мозга взрослого и новорожденного, размер мозга новорожденного, возможно, составлял 153–201 куб. См, аналогично тому, что предполагается для других австралопитеков.[14] Конфигурация мозга, по-видимому, была в основном подобна австралопитекам, но орбитофронтальная кора кажется более человечным.[15]
Общий, А. седиба анатомия черепа больше всего похожа на A. africanus. Однако MH1 имеет меньший череп, более широкий в поперечном направлении свод черепа, более наклонные в вертикальном направлении стенки теменная кость, и более широко расставленные височные линии. Так же, как Гомо, надбровная дуга менее выражена, скулы менее выпуклые, лицо не так сильно выступает (меньше прогнатизм ), есть небольшой подбородок.[1] Однако такие характеристики встречаются и у некоторых A. africanus черепа из Стеркфонтейн Член 4, который, по мнению Кимбел и Рак, мог указывать на то, что эти Гомо-подобные атрибуты были бы потеряны в зрелости. Кроме того, если прогнатизм измеряется с помощью передняя носовая ость вместо самого основания носа прогнатизм в MH1 попадает в диапазон, наблюдаемый в A. africanus.[2] Зубы у австралопитека довольно маленькие, и они больше соответствуют зубам ранних поколений. Гомо. Однако в отличие от Гомомоляры прогрессивно увеличиваются в размерах к задней части рта - в отличие от второго моляра, который является самым большим, - и бугорки расположены более близко друг к другу.[1]
Форма нижнечелюстная ветвь (стержень, соединяющий челюсть с черепом) сильно различается между MH1 и MH2. MH1 выше и шире; передняя и задняя граница почти вертикальны и параллельны, в отличие от непараллельных границ MH2 с вогнутой передней границей; и венечный отросток MH1 наклонен к спине с глубоким и асимметричным нижнечелюстная вырезка, тогда как MH2 имеет изогнутый венечный отросток с неглубокой нижней челюстью. По сравнению с моделями современных большие обезьяны, такие заметные различия превышают то, что можно было бы ожидать, если бы их можно было объяснить как следствие половой диморфизм или ювенильный статус MH1. Скелетно, А. седиба возможно, был сильно изменчивым видом.[16]
Торс
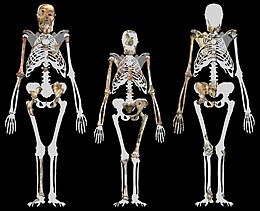
По оценкам, MH1 и MH2 были примерно одинакового размера, примерно 30–36 кг (66–79 фунтов). Это меньше, чем у многих современных гомининов, но приемлемо для австралопитека.[18] MH1 был около 130 см (4 фута 3 дюйма) в высоту, но он был подростком примерно с таким же развитием скелета, как у 12-летнего человеческого ребенка или 9-летнего шимпанзе. А. седиба, как и более ранние и современные гоминины, по-видимому, имели обезьяноподобную скорость роста, основанную на скорости развития зубов, поэтому MH1, возможно, достиг примерно 85% своего взрослого размера, принимая траекторию роста шимпанзе, или 80%, принимая траекторию, подобную человеческой. Это будет примерно 150 или 156 см (4 фута 11 дюймов или 5 футов 1 дюйм).[17]
MH1 пресервы 4 шея, 6 грудной, и 2 поясничный позвонки; и MH2 сохраняет 2 шейных, 7 грудных, 2 поясничных и 1 крестцовый позвонки.[19] Лордоз (человеческое искривление) и суставы шейных позвонков, указывающие на положение головы, подобное человеческому. Однако общая анатомия шейных позвонков похожа на обезьянью и указывает на гораздо более жесткую шею. А. седиба не хватает человечности плечевое сплетение (что указано в некоторых A. afarensis), а плечевое сплетение человека отвечает за нервную и мышечную иннервацию рук и кистей, улучшая двигательный контроль.[20] Как люди, А. седиба по всей видимости, имел гибкий поясничный ряд, состоящий из 5 позвонков - в отличие от 6 статичных позвонков у нечеловеческих обезьян - и демонстрирующий поясничный лордоз (искривление позвоночника у человека) соответствует обычной вертикальной позе. Тем не мение, А. седиба по всей видимости, у него очень подвижная поясница и усиленный поясничный лордоз,[19] которые могли быть задействованы в противодействии вращающим моментам, направленным внутрь при ходьбе гиперпронирующей походкой, предложенной для А. седиба.[21] MH1 сохраняет 2 верхних грудных, 1 среднегрудное и 3 нижних грудных ребра; и MH2 4 последовательных верхне-срединно-грудных и 3 нижних грудных ребра, соединенных с позвонками.[19] Это указывает на то, что А. седиба имел обезьяноподобно сжатую верхнюю часть груди, но анатомия таза, похожая на человеческую, может предполагать А. седиба имел широкую и человеческую нижнюю часть груди. Узкая верхняя часть груди затрудняла бы раскачивание руки во время ходьбы и ограничивала бы грудную клетку и предотвращала бы тяжелое дыхание и, следовательно, быструю ходьбу или бег на длинные дистанции. В отличие, А. седиба кажется, у него была узкая человеческая талия, переставленная внешние косые мышцы живота, и шире iliocostalis мышцы спины, которые улучшат эффективность ходьбы за счет противодействия боковому сгибанию туловища.[22]

Таз имеет несколько общих черт с ранним Гомо и Х. эргастер, а также KNM-ER 3228 от Кооби Фора, Кения и OH 28 из Олдувайское ущелье, Танзания, которые не относятся к какому-либо виду (хотя обычно классифицируются как Гомо spp.) Вдоль вертлужная впадина и крестец улучшение разгибания бедра, увеличение подвздошно-бедренная связка навесное оборудование, переносящее вес за центр вращения бедра, с большей опорой вдоль вертлужной впадины и подвздошная лопатка улучшение чередования наклон таза, и большее расстояние между вертлужной впадиной и седалищный бугорок сокращение момент рука на подколенные сухожилия. Возможно, это позволило сделать шаг вперед по-человечески А. седиба. Тазобедренный сустав, похоже, имел более человеческий образец нагрузки, чем H. habilis образец ОН 62.[1] В родовой канал из А. седиба кажется больше гинекоз (нормальное состояние человека), чем у других австралопитов, которые более платипеллоидны, хотя А. седиба не является полностью гинекозным, что может быть связано с меньшим размером мозга новорожденного (и, следовательно, головы). Как и у людей, родовые пути увеличились в диаметре в сагиттальном направлении (спереди назад), а лобковая кость загнута вверх.[14]
Верхние конечности
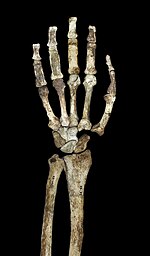
Как и другие австралопитеки и ранние Гомо, А. седиба имел несколько обезьяноподобных пропорций верхней части тела, относительно длинные руки, высокий плечевой указатель (от предплечья до плечевая кость соотношение) 84 и большие поверхности стыков. Обсуждается, является ли обезьяноподобная конфигурация верхних конечностей австралопитеков признаком древесного поведения или просто является базовой чертой, унаследованной от последнего общего предка больших обезьян, в отсутствие серьезного избирательного давления, чтобы принять анатомию руки, более похожую на человеческую. Плечи в положении пожатия плечами, лопатка имеет хорошо развитую подмышечная кайма и конический бугорок (важный для прикрепления мышц вокруг плечевого сустава) хорошо выражен.[1] Мышечные рубцы на ключице указывают на диапазон движений человека. Лопатка больше всего похожа на лопатку орангутаны с точки зрения размера гленоидная полость (который образует плечевой сустав) и его угол с позвоночником, хотя форма лопатки больше всего похожа на человеческую и шимпанзе. Плечевая кость имеет низкую степень перекручивания, в отличие от людей и африканских обезьян, что (вместе с короткой ключицей) предполагает, что лопатка была расположена дальше от средней линии, как у человека. Гомо, хотя он расположен выше на спине, как у других австралопитеков.[23] Обезьяньи качества рук, по-видимому, более выражены в А. седиба чем более древний A. afarensis, и если A. afarensis является предком А. седиба, это может указывать на адаптивный сдвиг в сторону древесного поведения.[24]
В локтевом суставе боковой и медиальный Надмыщелки плечевой кости удлиненные, как и у других австралопитеков и африканских обезьян, не являющихся людьми. Плечевая кость также имеет развитый гребень в локтевом суставе для поддержки брахиорадиалис мышца, сгибающая предплечье. Подобно нечеловеческим африканским обезьянам, существует сильная привязанность к бицепс на радиус и для трицепс на локтевая кость. Однако меньше механическое преимущество для бицепса и брахиалис.[23] Локтевая кость также поддерживает крепкое прикрепление локтевой сгибатель запястья. В локтевая ямка большой и глубокий, с выступающим трохлеарный киль, которые важны для поддержания устойчивости рук в вытянутом состоянии. Кости пальцев длинные, крепкий, и изогнутые, и сильные поверхностный сгибатель пальцев важно для сгибания пальцев.[1] Иногда их считают доказательством древесный поведение у австралопитеков. Рука также имеет относительно длинный большой палец и короткие пальцы, как и Гомо, что могло бы предложить точный захват важно при создании и использовании сложных каменные инструменты.[25]
Нижние конечности

Как и у других австралопитеков, голеностопный, коленный и тазобедренный суставы указывают на привычные двуногие. Кости ног очень похожи на кости A. afarensis. Лодыжка в основном похожа на человеческую, возможно, на человеческую. пяточное сухожилие.[26]
В таранная кость он толстый и больше похож на обезьян, не являющихся людьми, с изогнутой медиально шеей и низким углом скручивания шеи. Это обсуждается, если А. седиба имел человеческий свод стопы или если бы нога была более обезьяноподобной.[27] В пяточная кость наклонен под углом 45 градусов и заметно наклонен спереди назад, наиболее сильно у блока малоберцовой кости. Прочная блокировка малоберцовой кости указывает на сильную малоберцовые мышцы которые проходят через икры до щиколотки. На стопе отсутствует латеральный подошвенный бугорок (который может участвовать в рассеивании сил, когда пятка ударяется о землю при нормальной походке человека), что наблюдается у людей и A. afarensis.[1][26] Изящное тело пяточной кости и прочная лодыжка (костные выступы на каждой стороне лодыжки) весьма похожи на обезьяны, с менее эффективной передачей силы между пяточной костью и таранной костью и обезьяноподобной подвижностью в средней части стопы. А. седиба наиболее похоже на состояние, наблюдаемое у горилл, и нога могла быть функционально эквивалентна ноге A. africanus.[26][28]
Палеобиология
Рацион питания
Анализ фитолиты (микроскопические остатки растений) из зубной налет обоих образцов и анализ изотопов углерода показывает диету почти исключительно C3 лесные растения, несмотря на предположительно широкую доступность C4 растения в их смешанной среде саванны. Такой режим питания наблюдается и у современных шимпанзе саванны и предполагается для Ранний плиоцен Ardipithecus ramidus, но сильно отличается от любого другого раннего гоминина. Всего с двух зубов из MH1 было извлечено 38 фитолитов, 15 из которых соответствуют двудольные, 9 однодольные, а остальные 14 - неопределенные. Однодольные, вероятно, были получены из C3 травы и осока растет в хорошо орошаемых и тенистых местах, а другие фитолиты были получены из фруктов, листьев, древесины или коры. Хотя кору обычно едят другие приматы из-за ее высокого содержания белка и сахара, известно, что ни один другой гоминин не ел кору регулярно. Анализ стоматологических микропереносов аналогичным образом предполагает, что два гоминина Малапа ели твердую пищу, значения сложности варьируются от H. erectus и надежный P. robustus.[29] Тем не менее, челюсть, по-видимому, не была так хорошо приспособлена для производства высоких штаммов по сравнению с другими ранними гомининами, что может указывать на А. седиба не так сильно зависел от его способности обрабатывать механически сложные продукты питания.[30][31]
Интерпретация А. седиба как универсал травоядное животное C3 лесные растения согласуются с тем, что он хотя бы частично является древесным. Такая широкая диета могла позволить А. седиба занимали гораздо меньшие домашние участки, чем современные шимпанзе саванны, которые в основном потребляют только фрукты, так как А. седиба смог отказаться от коры и других устойчивых к изломам продуктов.[29]
Походка

Во время прогулки, А. седиба возможно, проявилось гиперпронирование голеностопного сустава, что привело к чрезмерному переносу веса внутрь во время фазы стойки. У современных людей-гиперпронаторов ступня сильно перевернута во время фазы замаха, и контакт с землей сначала осуществляется внешней границей ступни, что приводит к высокой крутящие моменты вращая всю ногу внутрь. Точно так же насадки для прямая мышца бедра и двуглавая мышца бедра мышцы в А. седиба соответствуют растяжениям, направленным по средней линии ног, бедер и колен. Этот способ ходьбы неидеален для современной анатомии человека, и гиперпронатеры имеют более высокий риск развития подошвенный фасциит, шины на голени, и большеберцовый стрессовые переломы. Чтобы противодействовать этому, А. седиба могли использовать подвижную среднюю часть стопы, а не жесткую, подобную человеку, среднюю часть стопы, что могло предотвратить чрезмерную нагрузку на лодыжку.[21]
Гиперпронирующая походка и связанный с ней набор адаптаций не были идентифицированы у других гомининов, и неясно, почему А. седиба разовьет это.[21] Подвижная средняя часть стопы также будет полезна при интенсивном лазании.[1][21][26] так что гиперпронация могла быть компромиссом между привычным двуногим мышлением и древесиной.[21]
Рождение

В вход в таз для женщины А. седиба оценивается, как 80,8 мм × 112,4 мм (3,18 дюйма × 4,43 дюйма) в длину и ширину (сагиттальный x поперечный), и, поскольку размер головы новорожденного оценивается как максимум 89,2 мм (3,51 дюйма), новорожденный, вероятно, вошел в вход в таз поперечно ориентирован, как и у других гомининов. Средняя плоскость входного отверстия таза сужена как минимум до 96,9 мм (3,81 дюйма), поэтому новорожденного, возможно, не нужно было поворачивать при рождении. Размеры входного отверстия таза были рассчитаны с помощью композитной реконструкции с участием молодого самца. седалищная кость; Точно так же родовой канал, возможно, был больше, чем рассчитывали. По оценкам, плечи были 74,3 мм (2,93 дюйма) в поперечнике, поэтому они не препятствовали бы родам больше, чем голова. Таким образом, новорожденный в точке наибольшего сужения занимал бы около 92,1% родовых путей, что давало бы достаточно места для полностью не вращающихся родов, как это имеет место у нечеловеческих обезьян и, возможно, других австралопитеков (хотя и полу- также предлагается ротационное рождение). Хотя можно пройти без вращения, средняя плоскость расширяется в переднезаднем направлении (спереди назад), и для новорожденного было бы больше места, если бы он вращался так, чтобы самая длинная длина головы совпадала с этим расширением.[32]
У современных людей, для сравнения, гораздо более трудоемкие и сложные роды, требующие полного вращения новорожденного, поскольку большой мозг и, следовательно, размер головы, а также жесткие плечи новорожденного человека значительно затрудняют прохождение через родовой канал. Используя оценку 145,8–180,4 куб. См для А. седиба размер мозга новорожденного, размер головы новорожденного был бы 73 мм × 89 мм (2,9 дюйма × 3,5 дюйма), как у новорожденного шимпанзе.[32]
Разработка
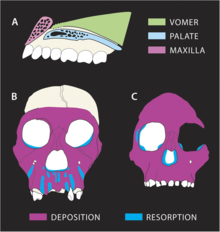
Траектория роста, по-видимому, заметно отличалась у MH1 от других гомининов. Назомаксиллярный комплекс (кость от носа до верхней губы) указывает на высокую степень резорбции кости, наиболее заметно в корни зубов передних зубов. Это контрастирует с A. africanus и A. afarensis которые являются депозитариями, отражая рост прогнатизма с возрастом. P. robustus также наблюдается резорбция верхней челюсти, но резорбция в MH1 распространяется вдоль передних зубов до собачья ямка возле скул, что приводит к мезогнатическому (несколько выступающему) лицу, в отличие от плоского лица в P. robustus. Поскольку резорбция происходит так близко к скуловым костям, это может объяснить, почему MH1 не дает выпуклых скул, характерных для A. africanus. Прорезывание зубов, вероятно, не повлияло на реконструкцию нижней части лица, поскольку у MH1 уже были все постоянные зубы. Тем не менее, меньший размер щечных зубов, возможно, позволил получить мезогнатическое лицо. Современный день оливковый колобус обезьяна могла иметь диету, похожую на А. седиба (что заметно контрастирует с типичными диетами ранних гомининов) и, по-видимому, имеет сходные модели роста лицевых костей, что может указывать на расходящиеся резорбции и образцы отложений в А. седиба от других гомининов отражает другие паттерны нагружения челюсти. Поля глазницы MH1 изогнуты, в то время как A. africanus, что может указывать на отложение костной ткани в А. седиба в регионах, где происходит резорбция костной ткани A. africanus.[33]
Патология
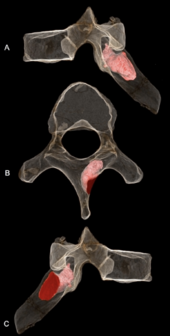
Правая пластинка шестого грудного позвонка MH1 представляет собой проникающую костная опухоль, наверное доброкачественный остеоид-остеома. Поражение проникает в глубину 6,7 мм (0,26 дюйма) и имеет ширину 5,9 мм (0,23 дюйма) и все еще оставалось активным на момент смерти. Он не проник в нервный канал поэтому, вероятно, это не вызвало каких-либо неврологических осложнений, и нет никаких доказательств сколиоз (аномальное искривление позвоночника). Это могло повлиять на движение лопатки и правого верхнего квадранта спины, возможно, вызвав острый или же хронический боль, мышечные расстройства или мышечные спазмы. Данный А. седиба может потребоваться способность лазать, расположение поражения рядом с местом введения трапеция, выпрямитель позвоночника, и ромбовидный мажор мышцы могут иметь ограниченные нормальные модели движения. MH1 имеет самый ранний диагностированный случай рак для гоминина по крайней мере на 200000 лет, что раньше SK 7923 возрастом 1,8-1,6 миллиона лет. плюсневой фрагмент представления остеосаркома из Сварткранс, Колыбель человечества. Опухоли в палеонтологической летописи гомининов встречаются редко, вероятно, из-за низкой заболеваемости в целом для приматов; ранние гоминины, вероятно, имели такую же заболеваемость, как и современные приматы. Развитие опухоли кости у ювенильного MH1 согласуется с общей тенденцией опухолей костей, чаще всего встречающихся у более молодых людей.[34]
MH1 и MH2 демонстрируют перимортальные (примерно в момент смерти) повреждения костей, соответствующие травма из-за применения грубой силы. Это согласуется с интерпретацией этого места как основания высокой шахты, действующей как естественная смертельная ловушка, в которую случайно попали животные. MH1 и MH2 могли упасть примерно с 5–10 м (16–33 футов) на наклонную груду гравия, песка и летучей мыши. гуано, что, вероятно, в какой-то степени смягчило падение. Для MH1 перимортальные переломы наиболее заметны на челюстной кости и зубах, хотя возможно, что эти травмы возникли в результате удара падающим предметом в дополнение к самому падению. MH2 имеет доказательства укрепляющий при травме с нагрузкой на предплечье и кисть и ударом в грудную клетку выявлен перимортальный перелом на правой стороне тела. Это первые смертельные случаи в летописи окаменелостей австралопитов, уверенно не связанные с хищничеством или естественными причинами.[35]
Палеоэкология

В 2010 году вместе с гомининами в фациях D и E было обнаружено в общей сложности 209 окаменелостей, не относящихся к гомининам, и таксоны определены из них: саблезубый кот Динофелис Barlowi, то леопард, то Африканская дикая кошка, то черноногий кот, то коричневая гиена, то накидка лиса, то мангусты Мезот Атилакса и Mungos, а генетика, Африканская дикая собака, а лошадь, а свинья, а клипспрингер, а Мегалотрагус антилопа, большая альселафина антилопа, родственник запряженный кустарник, родственник большое куду, а заяц.[4][36] Сегодня черноногий кот и накидка эндемичный в южноафриканские травы, кустарники и кустарники. Точно так же коричневая гиена населяет сухие открытые места обитания и никогда не наблюдалась в закрытых лесах. Динофелис и Атилаксс другой стороны, обычно являются индикаторами замкнутой влажной среды обитания. Это может указывать на то, что в этом районе была закрытая среда обитания, а также луга - судя по ареалу обитания лисицы, оба существовали в пределах 20 км.2 (7,7 квадратных миль) участка.[36]
В копролит плотоядного животного из фации D, содержащей пыльца и фитолиты Подокарпус или же Афрокарпус деревья, а также обломки древесины неустановленных хвойные породы и двудольные. Фитолиты злаков не обнаружены. В наши дни участок Малапа представляет собой пастбище, и Подокарпус и Афрокарпус находятся в 30 км (19 миль) от Афромонтане лес биом в каньонах 1500–1900 м (4 900–6 200 футов) над уровнем моря в Magaliesberg горный массив, где пожары встречаются реже. Это может указывать на то, что Малапа была более прохладной и влажной территорией, чем сегодня, что позволило снизить количество пожаров, чтобы позволить таким лесным растениям распространиться далеко за пределы естественных защищенных территорий. Малапа во время раннего плейстоцена, возможно, также находилась на несколько более низкой высоте, чем сегодня, с долинами и Магалисбергом, которые были менее выражены.[37]
Австралопитеки и ранние Гомо скорее всего предпочтительнее более прохладные условия, чем позже Гомо, поскольку нет стоянок австралопитеков, которые находились бы ниже 1000 м (3300 футов) над уровнем моря во время осаждения. Это означает, что, как и шимпанзе, они часто населяли районы со средней дневной температурой 25 ° C (77 ° F), опускающейся до 10 или 5 ° C (50 или 41 ° F) ночью.[38] Пещера Малапа в настоящее время находится на высоте 1442 м (4731 фут) над уровнем моря.[4] А. седиба жил рядом P. robustus и Х. эргастер/H. erectus. Потому что A. africanus вымерли примерно в это время, возможно, что Южная Африка была рефугиум за Австралопитек примерно 2 миллиона лет назад, когда началась большая климатическая изменчивость и изменчивость, и потенциально конкуренция с Гомо и Парантроп.[8]
Смотрите также
- Африканская археология
- Австралопитек африканский - Вымершие гоминиды из Южной Африки
- Homo ergaster - Вымершие виды или подвиды архаичного человека
- Homo gautengensis - Название предложено для вымершего вида гомининов из Южной Африки.
- Homo habilis - Архаичный человеческий вид от 2,1 до 1,5 млн лет назад
- Homo naledi - Южноафриканский архаичный человек с маленьким мозгом
- Paranthropus boisei - Вымершие виды гомининов Восточной Африки
- Paranthropus robustus - Вымершие виды гомининов Южной Африки
Рекомендации
- ^ а б c d е ж грамм час я j k л м п о Berger, L.R .; de Ruiter, D. J .; Черчилль, С. Э .; Schmid, P .; Карлсон, К. Дж .; Диркс, П. Х. Г. М .; Кибии, Дж. М. (2010). "Австралопитек седиба: новый вид Гомо-подобный австралопит из ЮАР ». Наука. 328 (5975): 195–204. CiteSeerX 10.1.1.729.7802. Дои:10.1126 / science.1184944. PMID 20378811. S2CID 14209370.
- ^ а б c d е Kimbel, W .; Рак, Ю. (2017). "Австралопитек седиба и появление Гомо: Сомнительные свидетельства из черепа молодого голотипа MH 1 ". Журнал эволюции человека. 107: 94–106. Дои:10.1016 / j.jhevol.2017.03.011. PMID 28526292.
- ^ Кинг, Дж. (4 июня 2010 г.). "Австралопитек седиба ископаемое, названное 17-летним студентом из Йоханнесбурга ". Центр истоков. Архивировано из оригинал 25 марта 2012 г.. Получено 9 июля 2011.
- ^ а б c d е Диркс, П. Х. Г. М .; Kibii, J.M .; Kuhn, B. F .; Steininger, C .; Черчилль, С. Э .; Kramers, J.D .; Пикеринг, Р .; Farber, D. L .; и другие. (2010). «Геологическая обстановка и возраст Австралопитек седиба из южной части Африки " (PDF). Наука. 328 (5975): 205–208. Дои:10.1126 / science.1184950. PMID 20378812. S2CID 206524717.
- ^ а б Пикеринг, Р .; Диркс, П. Х. Г. М .; Jinnah, Z .; и другие. (2011). "Австралопитек седиба 1,977 млн лет назад и влияние на происхождение рода Гомо". Наука. 333 (6048): 1421–1423. Дои:10.1126 / science.1203697. PMID 21903808. S2CID 22633702.
- ^ Irish, J. D .; Gautelli-Steinberg, D .; Legge, S. S .; и другие. (2013). «Стоматологическая морфология и филогенетическое« место »г. Австралопитек седиба". Наука. 340 (6129): 1233062. Дои:10.1126 / science.1233062. PMID 23580535. S2CID 206546794.
- ^ а б Вильмоар, В .; Kimbel, W. H .; Seyoum, C .; и другие. (2015). "Рано Гомо на 2,8 млн. лет от Леди-Герару, Афар, Эфиопия ". Наука. 347 (6228): 1352–1355. Дои:10.1126 / science.aaa1343. PMID 25739410.
- ^ а б Herries, A. I. R .; Martin, J.M .; и другие. (2020). "Современность Австралопитек, Парантроп, и рано человек прямоходящий в Южной Африке". Наука. 368 (6486): eaaw7293. Дои:10.1126 / science.aaw7293. HDL:11568/1040368. PMID 32241925. S2CID 214763272.
- ^ Балтер, Майкл (2010). «Кандидат в предки человека из Южной Африки вызывает похвалы и споры» (PDF). Наука. 328 (5975): 154–155. Дои:10.1126 / science.328.5975.154. PMID 20378782.
- ^ Черри, М. (8 апреля 2010 г.). «Утверждение о« предке человека »вызывает фурор». Природа. Новости природы. Дои:10.1038 / новости.2010.171.
- ^ Du, A .; Алемсегед, З. (2019). «Временные данные показывают, что Australopithecus sediba вряд ли является предком Homo». Наука. 5 (5): e9038. Дои:10.1126 / sciadv.aav9038. ЧВК 6506247. PMID 31086821.
- ^ а б Спур, Фред (5 октября 2011 г.). «Палеоантропология: Малапа и род Гомо". Природа. Дои:10.1038 / 478044a.
- ^ МакНалти, К. П. (2016). "Таксономия и филогения гомининов: что в имени?". Знания о естественном образовании. 7 (1): 2.
- ^ а б Kibii, J.M .; Черчилль, С. Э .; Schmid, P .; и другие. (2011). "Частичный таз Австралопитек седиба". Наука. 333 (6048): 1407–1411. Дои:10.1126 / science.1202521. PMID 21903805. S2CID 206532267.
- ^ Карлсон, К. Дж .; Стаут, Д .; Джашашвили, Т .; и другие. (2011). "Эндокаст MH1, Австралопитек седиба". Наука. 333 (6048): 1402–1407. Дои:10.1126 / science.1203922. PMID 21903804. S2CID 206533255.
- ^ Ritzman, T. B .; Terhune, C.E .; Gunz, P .; Робинсон, К. А. (2018). «Форма ветви нижней челюсти Австралопитек седиба предлагает единственный изменчивый вид ". Журнал эволюции человека. 100: 54–64. Дои:10.1016 / j.jhevol.2016.09.002. PMID 27765149.
- ^ а б Cameron, N .; Богин, Б .; Болтер, Д .; Бергер, Л. (2018). "Посткраниальное скелетное созревание Австралопитек седиба". Американский журнал физической антропологии. 163 (3): 633–640. Дои:10.1002 / ajpa.23234. PMID 28464269.
- ^ Холлидей, Т. У .; Черчилль, С. Э .; и другие. (2018). "Размер тела и пропорции Австралопитек седиба" (PDF). Палеоантропология: 406–422. Дои:10.4207 / PA.2018.ART118 (неактивно 16.09.2020).CS1 maint: DOI неактивен по состоянию на сентябрь 2020 г. (связь)
- ^ а б c Уильямс, С. А .; Острофский, К. Р .; и другие. (2013). "Позвоночный столб Австралопитек седиба". Наука. 340 (6129): 1232996. Дои:10.1126 / science.1232996. PMID 23580532. S2CID 206546736.
- ^ Meyer, M. R .; Уильямс, С. А .; Schmid, P .; Черчилль, С. Э .; Бергер, Л. (2017). "Шейный отдел позвоночника Австралопитек седиба". Журнал эволюции человека. 104: 32–49. Дои:10.1016 / j.jhevol.2017.01.001. PMID 28317555.
- ^ а б c d е DeSilva, J.M .; Holt, K. G .; Черчилль, С. Э .; и другие. (2013). «Нижняя конечность и механика ходьбы в Австралопитек седиба". Наука. 340 (6149): 1232999. Дои:10.1126 / science.1232999. PMID 23580534. S2CID 13288792.
- ^ Schmid, P .; Черчилль, С. Э .; Налла, С. (2013). «Морфология мозаики в грудной клетке Австралопитек седиба". Наука. 340 (6129): 1234598. Дои:10.1126 / science.1234598. PMID 23580537. S2CID 31073328.
- ^ а б Черчилль, С. Э .; Холлидей, Т. У .; Карлсон, К. Дж .; и другие. (2013). "Верхняя конечность Австралопитек седиба". Наука. 340 (6129): 1233477. Дои:10.1126 / science.1233477. PMID 23580536. S2CID 206547001.
- ^ Rein, T. R .; Харрисон, Т .; Карлсон, К. Дж .; Харвати, К. (2016). «Адаптация к задерживающему движению в Австралопитек седиба". Журнал эволюции человека. 104: 1–12. Дои:10.1016 / j.jhevol.2016.12.005. PMID 28317552.
- ^ Кивелл Т.Л., Кибии Дж. М., Черчилль С. Е., Шмид П., Бергер Л. Р. (2011). "Австралопитек седиба рука демонстрирует мозаики эволюции опорно-двигательного аппарата и манипулятивных способностей». Наука. 333 (6048): 1411–1417. Дои:10.1126 / science.1202625. PMID 21903806. S2CID 11610235.
- ^ а б c d Зипфель Б., ДеСильва Дж. М., Кидд Р. С., Карисон К. Дж., Черчилль С. Е., Бергер Л. Р. (2011). "Стопа и лодыжка Австралопитек седиба". Наука. 333 (6048): 1417–1420. Дои:10.1126 / science.1202703. PMID 21903807. S2CID 206532338.
- ^ Пранг, Т. К. (2015). «Положение задних конечностей Australopithecus sediba и эволюция продольной дуги гомининов». Научные отчеты. 5: 17677. Дои:10.1038 / srep17677. ЧВК 4667273. PMID 26628197.
- ^ Пранг, Т. К. (2016). «Подтаранный суставной комплекс Австралопитек седиба". Журнал эволюции человека. 90: 105–119. Дои:10.1016 / j.jhevol.2015.10.009. PMID 26767963.
- ^ а б Генри, Аманда Дж .; Ungar, Peter S .; Passey, Benjamin H .; Спонхеймер, Мэтт; Россоу, Ллойд; Бэмфорд, Мэрион; Сандберг, Пол; де Руйтер, Дэррил Дж .; Бергер, Ли (2012). "Диета Австралопитек седиба". Природа. 487 (7405): 90–93. Дои:10.1038 / природа11185. PMID 22763449. S2CID 205229276.
- ^ Ledogar, J. A .; Smith, A. L .; Benazzi, S .; и другие. (2016). "Механическое доказательство того, что Австралопитек седиба была ограничена в способности есть твердую пищу ". Nature Communications. 7 (10596): 10596. Дои:10.1038 / ncomms10596. ЧВК 4748115. PMID 26853550.
- ^ Daegling, D. J .; Карлсон, К. Дж .; Tafforeau, P .; de Ruiter, D. J .; Бергер, Л. (2016). «Сравнительная биомеханика нижних челюстей Australopithecus sediba». Журнал эволюции человека. 100: 73–86. Дои:10.1016 / j.jhevol.2016.08.006. PMID 27765151.
- ^ а б Лаудичина, Н. М .; Родригес, Ф .; ДеСильва, Дж. М. (2019). "Реконструкция рождения в Австралопитек седиба". PLOS ONE. 14 (9): e0221871. Дои:10.1371 / journal.pone.0221871. ЧВК 6750590. PMID 31532788.
- ^ Lacruz, R. S .; Bromage, T. G .; О'Хиггинс, П .; и другие. (2015). «Отчетливый рост носо-верхнечелюстного комплекса в Au. седиба". Научные отчеты. 5 (15175): 15175. Дои:10.1038 / srep15175. ЧВК 4606807. PMID 26469387.
- ^ Randolph-Quinney, P. S .; Уильямс, С. А .; Steyn, M .; и другие. (2016). «Остеогенная опухоль в Австралопитек седиба: Самые ранние доказательства неопластической болезни у гомининов ». Южноафриканский научный журнал. 112 (7–8). Дои:10.17159 / sajs.2016 / 20150470.
- ^ L'Abbé, E.N .; Symes, S.A .; Pokines, J. T .; Cabo, L. L .; и другие. (2015). «Свидетельства смертельных травм скелета у Malapa Hominins 1 и 2». Научные отчеты. 5 (15120): 15120. Дои:10.1038 / srep15120. ЧВК 4602312. PMID 26459912.
- ^ а б Kuhn, B. F .; Werdelin, L .; Hartstone-Rose, A .; Lacruz, R. S.; Berger, L. R. (2011). "Carnivoran Remains from the Malapa Hominin Site, South Africa". PLOS ONE. 6 (11): e26940. Дои:10.1371/journal.pone.0026940. ЧВК 3207828. PMID 22073222.
- ^ Bamford, M.; и другие. (2010). "Botanical remains from a coprolite from the Pleistocene hominin site of Malapa, Sterkfontein Valley, South Africa". Palaeontol. Afr. 45: 23–28.
- ^ Dávid-Barrett, T .; Данбар, Р. И. М. (2016). «Пересмотр двуногости и выпадения волос в эволюции человека: влияние высоты и расписания активности». Журнал эволюции человека. 94: 72–82. Дои:10.1016 / j.jhevol.2016.02.006. ЧВК 4874949. PMID 27178459.
дальнейшее чтение
- Уильямс, С. А .; Meyer, M. R .; Nalla, S.; и другие. (2018). "The Vertebrae, Ribs, and Sternum of Австралопитек седиба". Палеоантропология: 156–233. Дои:10.4207/PA.2018.ART113 (inactive 2020-09-16).CS1 maint: DOI неактивен по состоянию на сентябрь 2020 г. (связь)
- de Ruiter, D. J.; Churchill, S. E.; Berger, L. R. (2013). Рид, К. Э .; Fleagle, J.G .; Лики, Р. Э. (ред.). Australopithecus sediba from Malapa, South Africa. The Paleobiology of Australopithecus. Палеобиология и палеоантропология позвоночных. Springer Нидерланды. С. 147–160. Дои:10.1007/978-94-007-5919-0_9. ISBN 978-94-007-5919-0.
внешняя ссылка
- Реконструкции к Джон Гурч
- Skeletons Present an Exquisite Paleo-Puzzle на Наука
- What, if anything, is Австралопитек седиба? к Джон Д. Хоукс
- Malapa Hominin Site Entire Catalogue 2013 Part 1
- Malapa Hominin Site Entire Catalogue 2013 Part 2
- Human Timeline (Interactive) – Смитсоновский институт
| Таксономия (Hominins ) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Предки |
| ||||||||||||||||||||||||||
| Модели |
| ||||||||||||||||||||||||||
| Сроки | |||||||||||||||||||||||||||
| Другие |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| Места |  | ||||
|---|---|---|---|---|---|
| Окаменелости | |||||
| Антропология |
| ||||