Дманисские гоминины - Dmanisi hominins

В Дманисские гоминины,[1][2][3] Дманисцы[4] или Дманисский мужчина[5] были населением Ранний плейстоцен гоминины чьи окаменелости были обнаружены в Дманиси, Грузия. Возраст окаменелостей и каменных орудий, обнаруженных в Дманиси, составляет 1,85–1,77 миллиона лет.[6][7][8] делает гомининов Дманиси самыми ранними хорошо датированными окаменелостями гомининов в Евразия и наиболее хорошо сохранившиеся окаменелости ранних Гомо с одного сайта с сопоставимым временным контекстом. Хотя их точная классификация является спорной и спорной, окаменелость Дманиси весьма значима в исследованиях по ранние миграции гомининов из Африки поскольку они представляют собой старейшую летопись окаменелостей гомининов в Западной Евразии.[9] Гоминины Дманиси известны по более чем сотне посткраниальных окаменелостей и пяти знаменитым хорошо сохранившимся черепам, именуемым черепами Дманиси 1–5.
Хотя первоначально описывалась как популяция африканских видов Homo ergaster, или потенциально в раннем ответвлении более поздних азиатских H. erectus, таксономический статус гомининов Дманиси несколько неясен из-за их небольшого размера мозга, примитивной скелетной архитектуры и диапазона вариаций, наблюдаемых между черепами. Открытие массивной челюсти D2600 в 2000 году привело исследователей к гипотезе о том, что на этом участке присутствовало более одного таксона гомининов, и в 2002 году эта челюсть была определена как типовой образец нового вида. Homo georgicus. Более поздние анализы, проведенные исследовательской группой Дманиси, пришли к выводу, что все черепа, вероятно, представляют один и тот же таксон, хотя и демонстрируют значительные возрастные и половой диморфизм. Хотя исследователи Дманиси считают, что окаменелости могут быть заключены в H. erectus, либо как подвид Человек прямоходящий georgicus,[10] или под более популярным четырехчленом Homo erectus ergaster georgicus (просмотр Х. эргастер как подвид H. erectus и гоминины Дманиси как региональный вариант этого подвида),[11] это не универсальная точка зрения. Немного палеоантропологи все еще видишь имя Х. georgicus как более уместно, либо для некоторых,[12] или все окаменелости.[13]

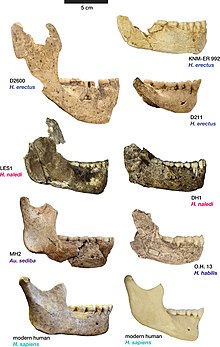
Анатомически гоминины Дманиси демонстрировали мозаику черт; обладающие некоторыми чертами, напоминающими более поздние и более производные H. erectus и современного человека, сохраняя при этом черты более ранних Гомо и Австралопитек. Длина и морфология их ног были в основном современными, и они были бы приспособлены к ходьбе и бегу на большие расстояния, но их руки, вероятно, были больше похожи на руки Австралопитек чем более поздние гоминины. Гоминины Дманиси также отличались бы от более поздних Гомо в их маленьком теле (145–166 см; 4,8–5,4 фута) и размере мозга (545–775 куб. см), оба из которых более сопоставимы с H. habilis чем позже H. erectus. Морфологические признаки, объединяющие все черепа, хотя степень их выраженности различается, включают крупные надбровные дуги и лица.
В плейстоцене климат Грузии был более влажным и заросшим лесами, чем сегодня, что сопоставимо с средиземноморский климат. Останки ископаемых в Дманиси были расположены недалеко от берега древнего озера, в окружении лесов и лугов, где обитала разнообразная фауна плейстоценовых животных. Благоприятный климат Дманиси мог служить убежищем для гомининов в раннем плейстоцене, и добраться до него можно было бы из Африки через Левантийский коридор. Найденные здесь каменные орудия относятся к Oldowan традиция, более примитивная, чем инструменты, сделанные и используемые более поздними H. erectus.
Таксономия
История исследований
Ранние раскопки в Дманиси

Дманиси расположен на юге Грузия, примерно в 85 километрах (52,8 мили) от столицы страны, Тбилиси. Он был основан как город в Средний возраст и, таким образом, был местом археологический интерес в течение некоторого времени, с известным местом археологических раскопок, расположенным в руинах старого города на мыс с видом на Машавера и Пиназаури реки.[14] Археологические раскопки начались в 1936 году по инициативе историка. Иване Джавахишвили, руководивший несколькими экспедициями. В 1982 году археологи в Дманиси обнаружили ямы глубиной 3 метра (10 футов), вырезанные в плотной песчаной глине. Археологи полагали, что ямы были созданы для каких-то экономических целей в средние века, и после их очистки на стенах и дне ям были обнаружены окаменелые кости животных. Грузинский палеобиологический институт Академии наук был незамедлительно проинформирован, и систематические палеонтологические раскопки начались в 1983 году, но закончились в 1991 году из-за финансовых проблем.[15]

Во время раскопок 1983–1991 гг. Было собрано большое количество окаменелостей животных, а также некоторые каменные орудия труда. Каменные орудия были быстро признаны в высшей степени архаичными, гораздо более примитивными, чем другие орудия, найденные в Восточной Европе. Биостратиграфически (датируемые путем сравнения с фауной на других хорошо датированных участках), орудия труда и окаменелости были датированы отложением в какой-то момент от позднего плиоцена до раннего плейстоцена.[15] Новые раскопки в Дманиси проводились с 1991 года, и к грузинским палеонтологам присоединились немецкие специалисты из Романо-германский музей в Кельне, Германия.[15] С 1991 по 1999 год экспедиции полностью финансировались Романо-германским музеем и проводились ежегодно.[15]
Открытие останков гоминина

Экспедиция 1991 года была высокопродуктивной, обнаружив многочисленные окаменелости животных и большое количество каменных орудий труда. Утром 25 сентября группа молодых археологов во главе с Медеей Ниорадзе и Антье Юстус обнаружила дугообразную кость.[15] Юстуса сразу заинтриговала необычная форма кости, и он интуитивно догадался, что это нижняя челюсть, что подтвердилось после того, как его раскопали более четко и обнажили зубы.[16]
Как отмечают руководители экспедиции, грузинские археологи и антропологи. Абесалом Векуа и Давид Лордкипанидзе (тогда в Тбилиси) были вызваны на место, и на следующее утро нижняя челюсть была освобождена от скалы вокруг нее, что заняло почти целый день. После освобождения нижняя челюсть безошибочно принадлежала примату и, что немаловажно, сохранила полный ряд зубов с небольшими признаками износа. Отсутствие износа предполагало, что примат был молодым, примерно 20–24 лет, хотя его классификация была пока неизвестна. После возвращения в Тбилиси нижнюю челюсть детально исследовали Векуа, Лордкипанидзе и археолог. Лео Габуниа. Было быстро решено, что он представляет гоминида, хотя его точное положение в семье было неясно. Хотя был обнаружен ряд примитивных особенностей, было ясно, что окаменелость (теперь получившая обозначение D211) была наиболее похожа на окаменелости Гомо, Не ранее австралопитеки. После продолжительного обсуждения Векуа и Габуния пришли к выводу, что гоминин Дманиси, вероятно, был одним из первых человек прямоходящий, и что это был самый ранний Гомо за пределами Африки. Это предположение и значение окаменелостей подтвердилось, когда было установлено, что возраст базальтов, лежащих непосредственно под плейстоценовыми отложениями, составляет около 1,8 миллиона лет.[16]
Раскопки на этом месте продолжались, хотя оказалось, что гоминины встречаются редко. В 1997 году правая плюсневая кость гоминина была обнаружена в том же слое, что и челюсть, что убедило ученых в том, что в Дманиси можно найти и другие останки гомининов. Дальнейшие открытия были сделаны в мае 1999 года. Из-за продолжительных периодов дождя место раскопок было повреждено. Археолог и участник экспедиции Гоча Киладзе обнаружил тонкую кость размером с монету и, поскольку она оказалась фрагментом черепа, привез ее в Тбилиси, где было установлено, что это фрагмент черепа приматов. Киладзе, Векуа, Лордкипанидзе, археолог Каха Кахиани и руководитель экспедиции 1999 г., археолог Георгий Копалиани, затем посетил сайт и обнаружил другие фрагменты. С помощью этих фрагментов они смогли соединить череп архаичного человека с обломанными зубами и отломанной верхней челюстью. В том же году был обнаружен более хорошо сохранившийся череп, и вместе эти два черепа позволили сделать выводы о природе и классификации ископаемых гомининов.[5] Первый череп, получивший название Череп 2 и получивший обозначение D2282, имел предполагаемую емкость черепа 650 куб. См, а второй череп, Череп 1 (D2280), имел расчетную вместимость 775 куб.[3] После изучения окаменелостей в течение почти года было установлено, что они несколько отличаются от H. erectus челюстями и черепами и были ближе к более ранним африканским видам Х. эргастер (теперь считается одним из первых африканских представителей H. erectus некоторыми). Обнаружение двух черепов было широко освещено в международных средствах массовой информации, и грузинские окаменелости впервые были широко признаны самыми ранними известными гомининами за пределами Африки.[5]
Дальнейшие открытия

Последовали новые открытия. В 2000 году была обнаружена еще одна челюсть гоминина (D2600),[3] на этот раз в несколько более низком слое (то есть старше), чем остальные окаменелости.[17] Эта челюсть была очень большой, с сильно развитыми задними коренными зубами. В следующем году был обнаружен череп 3 (D2700) и соответствующая ему челюсть (D2735), почти полностью сохранившиеся.[3] Из-за прорезывающихся зубов мудрости Череп 3 был определен как череп несовершеннолетнего.[4] В 2002 году был обнаружен беззубый череп старого человека, Череп 4 (D3444, соответствующая челюсть, D3900, была обнаружена в 2003 году). И Череп 3, и Череп 4 были отмечены как сохраняющие ряд очень примитивных характеристик. Последний череп, Череп 5 (D4500), был обнаружен в 2005 году. Череп соответствовал челюсти, найденной в 2000 году, и было установлено, что эти два черепа принадлежат одному человеку. Объем черепа 5 составляет 546 куб. См. Это самый низкий из всех черепов, и череп также является самым крепким из всех экземпляров.[3] Черепа примечательны не только своими уникальными особенностями. Череп 5 был первым полностью сохранившимся черепом взрослого гоминина раннего плейстоцена.[4] и Череп 4 - единственный беззубый гоминин, обнаруженный в таких ранних отложениях.[18]
Помимо черепов, обнаружено около сотни посткраниальных останков.[4] Первой обнаруженной посткраниальной окаменелостью была третья плюсневая кость, восстановлен в 1997 году.[19] Посткраниальные окаменелости включают кости всех частей тела, включая части рук,[20] ноги[21] осевой скелет (позвонки и ребра)[21] и ноги.[22] Кости, некоторые из которых уверенно связаны с Черепом 3, принадлежат как подросткам, так и взрослым людям.[20]
Вместе окаменелости в Дманиси представляют собой наиболее полную и богатую коллекцию ранних Гомо окаменелости в одном месте с сопоставимым временным контекстом. Различия в возрасте (например, череп 3 - младший взрослый, а череп 4 - значительно старше) и, предположительно, пол также дают уникальное представление о вариабельности ранних популяций Гомо.[4]
| Образ | Номер (а) черепа и образца | Емкость черепа | Обнаружил | Опубликовано | Заметки |
|---|---|---|---|---|---|
 | Дманиси Череп 1 D2280 | 775 куб.[3] | 1999[5] | 2000[23] | Тюбетейка взрослой особи.[24] Интерпретировано как мужчина из-за толстых надбровных дуг и других незначительных особенностей черепа.[25] |
 | Дманиси Череп 2 D2282 (нижняя челюсть D211) | 650 куб.[3] | 1999[5] (нижняя челюсть в 1991 г.)[16] | 2000[23] (нижняя челюсть в 1995 г.)[26] | Изящные черты лица интерпретируются как череп девушки-подростка.[24][25] |
 | Дманиси Череп 3 D2700 (нижняя челюсть D2735) | 600 куб.[27] | 2001[3] | 2002[28] | Череп молодой особи. Обычно морфология изящная, но верхняя клыки имеют крупные кроны и массивные корни; что затрудняет оценку его пола.[29] Некоторые черты характера позволяют предположить, что мужчина должен быть уместным.[25] |
 | Дманиси Череп 4 D3444 (нижняя челюсть D3900) | 625 куб.[30] | 2002[3] (нижняя челюсть в 2003 г.)[3] | 2005[31]/2006[32] | Череп пожилого человека, потерявшего все зубы, кроме одного.[33] Интерпретируется как мужчина.[34] |
 | Дманиси Череп 5 D4500 (нижняя челюсть D2600) | 546 куб.[3] | 2005[3] (нижняя челюсть в 2000 г.)[3] | 2013[35] (нижняя челюсть в 2002 г.)[36] | Череп взрослой особи. Череп 5 - первый найденный полностью сохранившийся череп взрослого гоминида раннего плейстоцена. Считается мужчиной из-за массивных и выдающихся черт черепа.[8][25] |
Классификация
Классификация гомининов Дманиси оспаривается, и ведется дискуссия о том, представляют ли они раннюю форму H. erectus, отдельный вид, названный Х. georgicus или что-то еще полностью продолжается.[2][13]
Ранние попытки классификации

Нижняя челюсть D211 была описана в 1995 году Габуниа и Векуа, которые сравнили ее с нижними челюстями других ранних Гомо; Х. эргастер, H. habilis и Х. рудольфенсис в Африке, H. erectus в Африке и Азии и архаичных Х. сапиенс в Европе. Из всех остальных Гомо, челюсть Дманиси была определена как наиболее похожая на окаменелости, приписываемые H. erectus в Африке, например ОН 22, с меньшим сходством с H. erectus окаменелости в Азии. Более близкое отношение к раннему Гомо такие как H. habilis и Х. рудольфенсис был исключен из-за того, что челюсти явно меньше, чем у этих видов, особенно под зубами на щеках. Размер зубов был отмечен как маленький, а с точки зрения морфологии зубы оказались особенно узкими, хотя зубы считались более или менее сопоставимыми с зубами в Х. эргастер/H. erectus, с наиболее сопоставимыми окаменелостями с точки зрения размера зубов OH 22, Чжоукоудянь черепа Мауэр 1, Тигениф 3, KNM ER 992 и KNM WT 15000.[37] По сравнению с этими другими образцами, D211 оказался наиболее отличительным в своем меньшем размере и уменьшении размера зубов. дистально (вдали от основной массы). Рассмотрев множество обнаруженных сходств, Габуния и Векуа пришли к выводу, что «наиболее разумная интерпретация этой челюсти состоит в том, что она принадлежала популяции H. erectus".[38]
Смесь примитивных и производных черт также была отмечена в анализе 1996 года палеоантропологами Гюнтером Бройером и Михаэлем Шульцем, которые сравнили нижнюю челюсть с ископаемыми нижними челюстями ранних времен. Гомо и Х. эргастер/H. erectus. В отличие от описания 1995 г., Бройер и Шульц пришли к выводу, что на основании наличия нескольких производных черт (обнаружение десяти основных черт, которые считаются аналогичными производным H. erectus), гоминин Дманиси, вероятно, был "прогрессивным" (т.е. производным, поздним) представителем H. erectus. Хотя они отметили, что такая классификация была бы сомнительной, поскольку возраст окаменелости 1,8 миллиона лет, они заявили, что челюсть все еще может представлять очень раннюю прогрессивную форму H. erectus.[39] В 1998 году палеоантропологи Антонио Росас и Хосе Бермудес де Кастро исследовали челюсть и обнаружили, что она демонстрирует уникальное сочетание особенностей, некоторые из которых, по отдельности, можно рассматривать как морфологические крайности внутри. Гомо. В то время как некоторые черты, такие как талонид (сокрушительная пята) премоляр зуб P4 был интерпретирован как более производный, многие из представленных особенностей, включая общую морфологию и размеры зубов и челюсти, считались примитивными. Поскольку похожая мозаика черт челюстей была обнаружена в африканских окаменелостях, Х. эргастер, Росас и Де Кастро предложили классифицировать челюсть как Гомо sp. индекс (aff. эргастер).[40]

Габуния и его коллеги описали черепа 1 и 2 в 2000 году и отметили, что, хотя лицевой скелет черепа 2 был фрагментарным, его предполагаемые пропорции и реконструированная морфология были очень похожи на образцы Х. эргастер восстановлен в Кооби Фора в Кении, и что зубной ряд, в частности, по размеру и морфологии напоминал зубной ряд, известный из Х. эргастер образцы, такие как KNM-WT 15000 и KNM ER 3733. Габуния и его коллеги отметили, что, хотя емкость черепа Дманиси была ниже, чем в среднем по Х. эргастер, черепа отличались от ранних Гомо такие как H. habilis и Х. рудольфенсис по ряду признаков, которые более тесно связывают окаменелости с Х. эргастер. Черты, отличающие гомининов Дманиси от ранних Гомо были отмечены как включающие хорошо развитые надбровная дуга, отсутствие гребень (иначе присутствует в начале Гомо и в других большие обезьяны ), большой орбиты, то премоляр зубы в верхняя челюсть одиночные корни и углы свод черепа. Были отмечены многочисленные черты, указывающие на тесную связь с Х. эргастер, включая наличие и морфологию надбровных дуг, общие пропорции лицевого скелета, относительную узость черепа за пределами лица (посторбитальное сужение ), а также сопоставимая высота свода черепа и толщина костей свода черепа. Те же функции, которые обычно используются для различения Х. эргастер из азиатских образцов H. erectus были обнаружены, чтобы отличить окаменелости Дманиси от азиатских H. erectus; особенно нижний свод черепа и несколько более тонкие кости свода черепа в H. erectus и меньшая емкость черепа у окаменелостей Дманиси. Было отмечено, что некоторые особенности присутствуют в окаменелостях Дманиси и азиатских H. erectus, но нет Х. эргастер, например, наличие супрамастоидный гребень ). Поскольку эти особенности также присутствовали в некоторых африканских окаменелостях, таких как Олдувайские гоминиды 9 и 12 считалось, что они не имели «особого филогенетического значения».[41] Габуния и его коллеги в заключение отнесли окаменелости Дманиси к Гомо напр. гр. эргастер ("напр. гр. эргастер"значение" группы, включающей эргастер") из-за комбинации особенностей, которые, по их мнению, выровняли окаменелости ближе к Х. эргастер чем раньше Гомо или азиатский H. erectus.[42] Габуния и его коллеги заявили, что комбинация особенностей делает возможным то, что гоминины Дманиси были предшественниками обоих более поздних H. erectus в Азии и гоминины, предки Х. сапиенс.[43]
Классификация после открытия других окаменелостей

В 2002 году Векуа и его коллеги описали череп 3 (D2700), включая связанную с ним нижнюю челюсть (D2735). Путем сравнения с другими окаменелостями гомининов со всего мира Озеро Туркана в Кении было установлено, что череп имеет ряд анатомических особенностей с черепами Х. эргастер/H. erectus, например, форма плоскоклеточная часть височной кости, то скуловой (скулы) корень очень толстый, а вдоль сагиттальный средняя линия. Это означает, что, несмотря на то, что череп 3-го черепа исключительно мал (объем черепа 600 куб. См, что близко к среднему значению). H. habilis), а его лицо очень похоже на лицо KNM ER 1813 (экземпляр H. habilis), особенно в профиль, Skull 3 больше похож на маленькую H. erectus чем H. habilis. Несмотря на некоторые различия, D2700 был признан в целом похожим на черепа 1 и 2, его нижняя челюсть напоминала нижнюю челюсть D211, и Векуа и его коллеги рассматривали все окаменелости как принадлежащие к одному и тому же таксону, не видя достаточных оснований для отнесения их к нескольким видам. . Череп 3 был временно назван человек прямоходящий (=эргастер). Исследователи также кратко упомянули нижнюю челюсть D2600 (на данный момент еще не описанную), чтобы подчеркнуть тот факт, что некоторые из окаменелостей гомининов, обнаруженных в Дманиси, отличаются от типичной морфологии H. erectus, но тот Череп 3, тем не менее, можно было рассматривать как «чрезвычайно мелкомозгового представителя этого вида».[27]
Нижняя челюсть D2600 была также описана в 2002 году Габуниа, Векуа и Лордкипанидзе вместе с французскими археологами и палеоантропологами. Генри де Ламли и Мари-Антионетт де Ламли.[24] До открытия нижней челюсти гоминины Дманиси были классифицированы как Х. эргастер или H. erectus казалась относительно чистой, но D2600 отличался своими большими размерами, морфологическими особенностями и пропорциями зубов не только от ранее обнаруженной челюсти в Дманиси, но также и от всех других челюстей гомининов, обнаруженных на сегодняшний день, сочетая примитивные черты Австралопитек и рано Гомо с производными функциями, иначе видимыми в H. erectus.[18] Габуния и его коллеги считали различия в размерах и пропорциях не только H. erectus и Х. эргастер, но и H. habilis и Х. рудольфенсис быть достаточным для создания нового вида, который они назвали Homo georgicus. D2600 был обозначен как типовой образец Х. georgicus, и все ранее обнаруженные останки гомининов были отнесены к этому новому виду.[24] Хотя черепа 1 и 2 были гораздо более изящными, чем новая крепкая нижняя челюсть, исследователи определили, что это был пример выраженного полового диморфизма только у одного вида, полагая, что изящные окаменелости представляют собой женщин. Габуния и его коллеги переводили Х. georgicus как потомок H. habilis или Х. рудольфенсис и ранний вид "у корней Гомо ветвь "," предсказывая появление Homo ergaster".[44] Палеоантрополог Санг-Хи Ли поддержал классификацию всех окаменелостей гомининов Дманиси как принадлежащих к одному и тому же виду (хотя и не прокомментировал, должен ли этот вид относиться к одному виду). H. erectus или Х. georgicus) в 2005 году, отметив, что, несмотря на различия в емкости мозга между черепами, они не были более морфологически отличны друг от друга, чем особи разных полов современных человекообразных обезьян.[45]

Лордкипанидзе и его коллеги описали череп 4 и его нижнюю челюсть в 2006 году, отметив, что он был похож на окаменелости, обнаруженные ранее, и заявив, что, за возможным исключением нижней челюсти D2600, все окаменелости Дманиси можно отнести к одному виду. Исследователи отметили, что популяция гомининов имеет несколько общих черт с более примитивными гомининами, такими как Австралопитек и раньше Гомо, особенно низкий объем черепа, но многие другие аспекты их морфологии согласовывали их с H. erectus, особенно африканские H. erectus (Х. эргастер), хотя они обладают некоторыми чертами, которые иначе наблюдаются только у азиатских H. erectus. На анатомической основе они утверждали, что гоминины Дманиси были близки к «стеблю» H. erectus, с прочими H. erectus группы являются более производными, и что грузинское население потенциально могло быть предком более поздних ветвей этого вида в Азии.[46] В том же году сравнительный анализ черепов с 1 по 4 и нижней челюсти D2600 палеоантропологом Дж. Филипом Райтмайром, Лордкипанидзе и Векуа снова пришел к выводу, что черепа с 1 по 4 можно отнести к одному и тому же виду, но что статус D2600 был более сомнительным. .[47] Они отметили, что хотя окаменелости были похожи на H. habilis в некоторых отношениях, особенно по размеру и (для некоторых) черепной емкости, у них было гораздо больше общих черт с H. erectus. В этом отношении многие из примитивных функций можно было бы просто интерпретировать как примитивные удержания. Райтмайр, Лордкипанидзе и Векуа пришли к выводу, что если некоторые из H. habilis-подобные черты, такие как размер, емкость черепа и части морфологии лица, были рассмотрены плезиоморфный и примитивных ретенций, нет причин исключать черепа с 1 по 4 из H. erectus. Из-за некоторых уникальных черт и этих примитивных ретенций Райтмайр, Лордкипанидзе и Векуа предположили, что гоминины Дманиси представляют собой ранний подвид H. erectus.[10] Хотя другие были не уверены, Векуа поддержал классификацию D2600 как представителя особого вида, отдельного от остальных окаменелостей, предпочитая сохранить его обозначение как Х. georgicus.[48] Они отметили, что если будущий анализ покажет, что D2600 принадлежал к той же популяции гомининов, что и другие окаменелости, то обозначение подвида было бы соответствующим образом Человек прямоходящий georgicus, но если бы он был отличным (как Х. georgicus), для других окаменелостей необходимо будет выбрать новое название подвида.[10]

Сравнительный анализ D211 и D2600 в 2006 г., проведенный палеоантропологами Мэтью М. Скиннером, Адамом Д. Гордоном и Николь Дж. Коллард, обнаружил, что степень диморфизма, выраженного между двумя нижними челюстями, была больше, чем ожидалось у современных человекообразных обезьян и человека, а также у человека. другие вымершие виды гомининов. Поскольку диморфизм оказался значительно больше, чем ожидается в таких родах, как Сковорода, Горилла и Гомо, Скиннер, Гордон и Коллард предложили две альтернативные гипотезы; либо окаменелости представляли один таксон с необычно высоким половым диморфизмом, включение которого в Гомо было сомнительно, или то, что D2600 следует рассматривать как представителя отдельного, второго вида гомининов (т.е. Х. georgicus).[49] Более подробный сравнительный анализ нижних челюстей в 2008 году с учетом большего количества анатомических особенностей, проведенный Райтмайром, Лордкипанидзе и палеоантропологом Адамом Ван Арсдейлом, пришел к выводу, что, хотя диморфизм между нижними челюстями был чрезмерным по сравнению с современными людьми, а некоторые шимпанзе, это было сравнимо (или, в некоторых случаях, меньше) с диморфизмом между гориллами. Они пришли к выводу, что «на наш взгляд, в настоящее время нет убедительных анатомических оснований для классификации каких-либо окаменелостей Дманиси, кроме одного вида», но отметили, что этот вид обладал бы более значительным половым диморфизмом, чем более поздний Гомо.[50] Предпочитая обозначение H. erectus, исследователи отметили, что хотя H. erectus Обычно считается, что это не диаморфизм, некоторые окаменелости, такие как меньшие черепа, найденные в Илерет и Олоргесайли в Кении и черепа большего размера, найденные в Олдувайское ущелье, Танзания и Бури Эфиопия, могла опровергнуть это мнение.[51]
Анализ зубов черепов 2 и 3 и нижней челюсти D2600, проведенный в 2008 году Лордкипанидзе, Векуа и палеоантропологами Марией Мартинон-Торрес, Хосе Марией Бермудес де Кастро, Аидой Гомес-Роблес, Анн Мергвелашвили и Лейре Прадо, показал, что, как и другие части окаменелостей , зубы тоже показали сочетание примитивных Австралопитек- и H. habilisчерты типа и многое другое, производное H. erectusчерты типа. Зубы черепов 2 и 3 оказались похожими, тогда как D2600 несколько различались по размеру зубов и морфологии корней. Однако это не обязательно само по себе указывает на то, что D2600 представляет другую популяцию или вид, поскольку такие же различия в размерах зубов наблюдаются в окаменелостях, упомянутых H. habilis.[52] Палеоантрополог П. Джеймс Макалузо мл.пришли к такому же выводу в анализе 2010 года, заключив, что черепа 2 и 3 могут быть отнесены к одному и тому же виду, но менее ясно, можно ли отнести D2600 к тому же виду, что и остальные.[53]
Классификация после описания Черепа 5

Череп 5, обнаруженный в 2005 году и описанный в 2013 году Лордкипанидзе и его коллегами, после его описания был определен как принадлежащий тому же человеку, что и нижняя челюсть D2600, и вместе эти две окаменелости значительно расширили морфологический диапазон окаменелостей гомининов Дманиси.[8] Лордкипанидзе и его коллеги интерпретировали череп 5 как часть той же популяции, что и остальные окаменелости Дманиси. Этот вывод был сделан, поскольку все окаменелости гомининов Дманиси происходят из одного и того же пространственного и временного контекста, а также потому, что новый анализ ископаемого материала показал, что черепа Дманиси находились в пределах диапазона вариаций, показанных у шимпанзе (Пан троглодиты троглодиты, П. т. Verus, и П. т. Schweinfurthii ), бонобо (P. paniscus) и современные образцы человека. Межвидовые различия во всех этих выборках были в основном из-за различий в размере и ориентации лица по отношению к мозговой коробке, оба различия также выражены в образце Дманиси. Лордкипанидзе и его коллеги интерпретировали, что маленькие и более ортогнатические черепа представляли женщин и / или подростков, а более прогнатические черепа и черепа с большим лицом - мужчин.[54] Большая степень изменчивости, выраженная в окаменелостях Дманиси, привела Лордкипанидзе и его коллег к предположению, что вариации, наблюдаемые в других окаменелостях гоминид плиоцена и плейстоцена, обычно используемые для обоснования нескольких различных ископаемых видов, могли быть неверно истолкованы как разнообразие видов. Таким образом, морфологическое разнообразие современных африканских гомининов, обычно используемое для оправдания Х. эргастер как вид, отличный от H. erectus, таким образом, вместо этого может быть связано с региональными различиями в одной развивающейся линии гомининов (H. erectus). Имея это в виду, африканский материал классифицируется как H. erectus ergaster (а хроноподвиды а не отдельный вид), и поскольку считается, что гоминины Дманиси произошли от ранней миграции H. erectus происхождением из Африки, было определено, что они лучше всего подходят для H. e. эргастер с четырехчленом (4-частным) названием; H. e. е. georgicus. Исследователи посчитали возможным, что ранее Гомо, такие как H. habilis и Х. рудольфенсис также принадлежали к той же единой развивающейся линии Гомо, хотя никаких морфологических сравнений для проверки этой теории не проводилось.[55]
Палеоантропологи Джеффри Х. Шварц, Ян Таттерсалл и Чжан Чи ответили на исследование 2013 года в 2014 году, не согласившись с идеей, что все пять черепов принадлежали к одному виду. Шварц, Таттерсалл и Чи также предложили использовать такие подвиды, как H. e. е. georgicus, с квадриномным названием, не использовался в зоологической номенклатуре. Что наиболее важно, Шварц, Таттерсалл и Чи задались вопросом, были ли морфологические сравнения достаточно подробными, чтобы прийти к такому выводу, и поставили под сомнение методы, которые Лордкипанидзе и его коллеги использовали для определения того, что является межвидовой изменчивостью, а что нет. Исследователи не усмотрели того факта, что окаменелости были из одного и того же места, и относительно короткого периода времени, достаточного для определения того, что все они произошли от одного вида и что предыдущие утверждения Гориллавариация нижней челюсти, но Х. сапиенс/Сковородавариации черепа не могут быть правильными одновременно. Они также задались вопросом, действительно ли все морфологические различия могут быть связаны с возрастом, износом и патологией. Некоторые черты черепов и зубов всех черепов Дманиси были выдвинуты как «потенциально отличительные для вида особенности», и Шварц, Таттерсолл и Чи пришли к выводу, что по крайней мере нижняя челюсть D2600 и, следовательно, череп 5 в целом должны оставаться классифицированными как отдельные виды, Х. georgicus, написав, что «отрицать особую идентичность этого гоминина - значит отрицать полезность морфологии в систематике - радикальное предложение, под которым немногие согласились бы».[12]
Исследовательская группа Дманиси ответила Шварцу, Таттерсоллу и Чи в том же году, утверждая, что окаменелости представляют один вид. Они отметили, что различие Х. georgicusи дальнейшее предположение о том, что некоторые из других черепов также могут представлять отдельные таксоны, означало бы, что Дманиси был домом по крайней мере для четырех различных таксонов гоминид и, таким образом, «удерживал мировой рекорд по разнообразию палеовидов гоминидов, документально подтвержденный в одном месте, которое простирается на площади всего 40 м² и, вероятно, занимает всего пару столетий ". Команда Дманиси написала, что Шварц, Таттерсалл и Чи сознательно проигнорировали предыдущий морфологический анализ, а также отметили, что изменение состояния характера у азиатских и африканских народов. Гомо образцы и окаменелости Дманиси предполагают, что окаменелость не может быть отнесена к разным видам, обвиняя Шварца, Ттаерсалла и Чи в эффективном отрицании морфологических свидетельств окаменелостей Дманиси, которые не соответствовали их гипотезе. Одна из основных отличительных черт, отмеченных Шварцем, Таттерсоллом и Чи, количество корней премоляров, была отмечена как не имеющая таксономического значения, поскольку современные люди к югу от Сахары демонстрируют значительные различия в этой конкретной особенности. Название Homo erectus ergaster georgicus был также защищен тем, что он использовался для обозначения местной популяции подвида, подобно тому, как квадриномы используются в ботаника. Исследователи отметили, что хотя использование квадриномов не регулируется Международный кодекс зоологической номенклатуры, это не считается недействительным.[11]
Анализ черепа 5, проведенный в 2017 году, в сравнении с другими черепами и черепами Х. сапиенс, Paranthropus boisei и других архаичных гомининов, группа подтвердила, что вариации между окаменелостями Дманиси не были чрезмерными по сравнению с вариациями у большинства других гомининов, с некоторыми особенностями, такими как определенные размеры средней части лица, даже более вариабельными у современных людей.[56] Хотя определенные черты были отмечены как устанавливающие череп 5 «на периферию распределения форм Дманиси», они пришли к выводу, что «ни эти различия, ни пропорции нижней челюсти D2600 не дают достаточных оснований для обозначения черепа 5 как« голотип морфологически ». очень самобытный вид Х. georgicus'». Результаты анализа, в котором черепа сравнивались со многими экземплярами обоих H. erectus и H. habilis несколько поставил под сомнение нынешнее признание разнообразия видов на раннем этапе Гомо до сих пор было обнаружено, что гоминины Дманиси имеют много общего с обоими видами. Исследователи установили, что гоминины Дманиси "тоже нельзя однозначно отнести к H. habilis или чтобы H. erectus"и что там, что касается раннего Гомо, был «континуумом форм»; Череп 5 имеет много общих черт с H. habilis тогда как Череп 1 с самым большим мозгом больше похож на африканский Х. эргастер/H. erectus.[57] Это привело исследователей к предположению, что H. erectus и H. habilis составляют единую эволюционную линию, возникшую в Африке, а затем распространившуюся по всей Евразии. Филогенетически популяция Дманиси была предложена как часть анагенетический последовательность, произошедшая от H. habilis и наследственный к более позднему H. erectus, размещенный у основания H. erectus происхождение и уже дифференцировано от H. habilis.[58] Хотя они представлены всего несколькими окаменелостями, морфология Х. рудольфенсис был определен как уникальный, предполагая, что Х. рудольфенсис был отдельным видом, частью отдельной эволюционной линии.[59]
Хронология и география

Время первой архаической миграции человека из Африки и идентичность вида гомининов, предпринявших эту миграцию, спорны. Это происходит из редких окаменелостей гомининов раннего плейстоцена за пределами Африки. До открытия черепов Дманиси самые ранние известные окаменелости гомининов в Европе и Азии были либо слишком неполными и фрагментарными, чтобы их можно было надежно идентифицировать на уровне видов, либо имели морфологические черты, характерные для региона, где они были обнаружены. Более того, большинство участков, где были обнаружены эти окаменелости, были восстановлены в сохраненном геологическом контексте, который можно было надежно датировать. Из-за этого возникли споры относительно того, распространились ли архаические люди из Африки в позднем плиоцене или раннем плейстоцене в результате сети экоморфологических факторов или около 1 миллиона лет назад в результате технологических инноваций, таких как Ашельский инструментальная культура.[14][60]
Гоминины Дманиси, датированные представлением самых ранних известных гомининов в Европе.[61] Отложения плейстоцена в Дманиси отложены непосредственно на толстом слое вулканической породы, которая была радиометрически датированный до 1,85 миллиона лет. Контуры отложений плейстоцена показывают, что между отложением этих вулканических пород и отложением более новых отложений прошло относительно мало времени. Через палеомагнитный анализа было определено, что отложениям, вероятно, около 1,77 миллиона лет, в то время, когда магнитная полярность планеты была изменена. Окаменелости других животных, найденных на этом месте, например, грызунов. Мимомис (который, как известно, жил только 2,0–1,6 миллиона лет назад), подтверждает эту дату.[4]
В 2010 году уровень содержания гомининов в ископаемом Дманиси был датирован через аргон-аргонное датирование возрастом 1,81 ± 0,03 миллиона лет, лишь немного моложе нижележащего слоя вулканической породы. Эта более ранняя дата противоречила предыдущей оценке возрастом 1,77 миллиона лет, основанной на палеомагнитных данных. Поскольку челюсть D2600 была обнаружена в несколько более низком слое, считалось возможным, что эта конкретная окаменелость была еще более раннего возраста, но поскольку не было оценок скорости осаждения на этом участке, также могло быть только несколько тысячелетий, разделяющих челюсть от остальных окаменелостей.[61] Каменные орудия, найденные на стоянке Дманиси, имеют возраст от 1,85 миллиона до 1,78 миллиона лет, что позволяет предположить, что гоминины населяли это место на протяжении всего времени между двумя предполагаемыми возрастами самих окаменелостей.[7] Помимо окаменелостей Дманиси, на острове были обнаружены каменные орудия труда, изготовленные гомининами. Лессовое плато в Китае и датируется 2,12 миллионами лет, а это означает, что гоминины должны были покинуть Африку до этого времени.[62]
В позднем плиоцене и раннем плейстоцене Грузия могла служить убежищем для групп гомининов, живущих в регионах с истощенными ресурсами. Окружающая среда в Дманиси была бы благоприятной для гомининов из-за физической географии региона, подходящих биомов и того факта, что Большой Кавказ горный массив служил преградой для воздушных масс с севера. Гоминины, вероятно, достигли Грузии через Левантийский коридор, который уже существовал в это время. Гоминины, возможно, обосновались в Дманиси до того, как расширились в других местах, поскольку окаменелости животных того же возраста присутствуют в местах в Румынии, на Балканах и даже в Испании, некоторые из них сопровождаются каменными орудиями, напоминающими найденные в Дманиси.[63]
Анатомия
Череп

Объем черепа гомининов Дманиси колеблется от 546 до 775 куб.[4] в среднем 631 куб.[64] Таким образом, размер их мозга совпадает с размером мозга H. habilis (c. 548–680 куб. См) и ниже стандартной черепной емкости, которая в противном случае приписывалась бы H. erectus (800–1000 куб.[4]). В коэффициент энцефализации (мера размера мозга по отношению к размеру тела) гомининов Дманиси (на основе черепов с 1 по 4) находится в диапазоне 2,6–3,1, что соответствует нижнему пределу оценок для Х. эргастер/H. erectus и больше похоже на H. habilis и австралопитеки.[22] Коэффициент энцефализации черепа 5 оценивается в 2,4, в пределах диапазона вариации для Австралопитек.[65]
Единственный полностью завершенный череп, найденный в Дманиси, - это Череп 5, который можно отличить от всех других известных окаменелостей. Гомо экземпляров большим прогнатическим лицом и маленькой черепной коробкой.[65] Сочетание больших зубов и большого лица с маленькой черепной коробкой было неизвестно в раннем возрасте. Гомо,[2] и эти два признака ранее отдельно использовались для определения разных видов. Если бы черепная коробка и лицо черепа 5 были обнаружены как отдельные окаменелости в разных местах, вероятно, они были бы отнесены к разным видам.[66] Несмотря на внешнее морфологическое сходство с более ранними Гомо, анатомия его черепной коробки значительно больше похожа на более поздние виды, такие как H. erectus.[2]
Череп 5 несколько отличается от других индивидов Дманиси, с более выдающимися и массивными черепными структурами (то есть лицом и челюстью).[8] Это не обязательно означает, что Череп 5 представляет другую популяцию гомининов; два других более или менее полных черепа, Черепа 3 и 4, принадлежат подростку (и, следовательно, не полностью развиты) и пожилому человеку, соответственно. Череп 5 указывает на то, что маленький мозг, большие лица и в целом прогнатическая и прочная морфология находились в пределах диапазона вариаций популяции гомининов Дманиси. Судя по черепам и посткраниальному материалу, гоминины Дманиси, по-видимому, были людьми с маленьким мозгом, с ростом, массой тела и пропорциями конечностей на нижней границе диапазона современных человеческих вариаций.[65]
Несмотря на очевидные различия между черепами, многие черты у них общие. Все они имеют относительно небольшой размер мозга, и надбровные дуги видны у всех взрослых особей, только менее заметны у субзарядного черепа 3. Хотя он наиболее массивен в черепе 5, лицо относительно прогнатическое и массивное на всех участках. черепа.[67]
Посткраниальная анатомия
До открытия окаменелостей Дманиси знания посткраниальной морфологии в раннем Гомо был очень ограничен. Хорошо сохранившиеся окаменелости более ранних гомининов, таких как Австралопитек и позже Гомо, например, хорошо сохранившийся скелет KNM WT 15000 («Мальчик Туркана»; 1,55 миллиона лет назад Х. эргастер/H. erectus), дал мало информации о ранних изменениях в пропорциях тела и росте. Австралопитек были небольшими, около 105 см (3,4 фута) в высоту, и имели промежуточные пропорции конечностей между современными людьми и другими человекообразными обезьянами, в то время как пропорции тела и рост мальчика Туркана были более или менее современными. Посткраниальные окаменелости, приписываемые H. habilis и Х. рудольфенсис фрагментарны, поэтому время и средства перехода от гомининов, способных к двуногому поведению (Австралопитек) гомининам, которые были обязательно двуногими (Х. эргастер) осталось неясным. В этом отношении окаменелости Дманиси заполняют ряд пробелов.[22]
Посредством расчетов, основанных на размере их костей (полный скелет еще не восстановлен), люди Дманиси были приблизительно 145–166 см (4,8–5,4 футов) в высоту и весили около 40–50 кг (88–110 фунтов). Они были меньше, чем Х. эргастер в Африке, возможно, из-за того, что они более примитивны (H. habilis также был меньше, чем Х. эргастер) или из-за адаптации к другому палеоэкологическому контексту. Пропорции конечностей (измеряются по длине бедренная кость относительно большеберцовая кость ) в окаменелостях Дманиси сопоставимы с окаменелостями современных людей, но также сопоставимы с некоторыми из самых ранних Гомо и окаменелости, упомянутые Австралопитек гархи возрастом 2,5 миллиона лет. По абсолютной длине ног гоминины Дманиси были больше похожи на более поздних Гомо (включая современных людей), чем австралопитеки,[22] хотя длина ног и морфология плюсневых костей у гомининов Дманиси не были так получены, как более поздние Х. эргастер/H. erectus (например, KNM WT 15000). Это может указывать на то, что развитие улучшенных характеристик ходьбы и бега было не внезапным изменением, а непрерывным процессом на протяжении раннего и среднего плейстоцена.[68]
Перекрут плечевой кости (угол между проксимальный и дистальный суставная ось плечевая кость ) влияет на диапазон движений и ориентацию рук относительно торс. У современных людей лопатка (который в противном случае может ограничить движение) помещается дорсально, что компенсируется высокой степенью перекрута плечевой кости. Для сравнения, скручивание в окаменелостях Дманиси довольно низкое, что указывает на различное движение и ориентацию рук. Это могло означать, что руки обычно были ориентированы более лежачий (по горизонтали) и что плечевой пояс мог бы быть расположен больше сбоку. Спортсмены, которым требуется высокий уровень подвижности рук, как правило, имеют меньшее перекручивание плечевой кости, и, таким образом, гоминины Дманиси могли бы обладать разнообразными движениями рук. Перекрут плечевой кости также низкий (или полностью отсутствует) у H. floresiensis, что означает, что это может быть основной чертой Гомо. В любом случае, функциональность и морфология рук у гомининов Дманиси, похоже, были больше похожи на руки более ранних Гомо или австралопитеки, чем современные люди.[69]
В целом позвоночник у гомининов Дманиси, похоже, были больше похожи на колючки современных людей и H. erectus чем к шипам австралопитеков. Ископаемые позвонки, найденные на выставке в Дманиси поясничный лордоз, ориентация фасеточные суставы предполагает, что диапазон сгибания позвоночника у гомининов Дманиси был сопоставим с современными людьми, а относительно большие площади поперечного сечения позвонков указывают на устойчивость к повышенным сжимающим нагрузкам, предполагая, что гоминины были способны бегать и ходить на большие расстояния. Поскольку были найдены окаменелости голени, от бедренной кости до плюсневых костей, можно восстановить ориентацию и положение ступней гомининов Дманиси относительно направления их ходьбы. У гомининов Дманиси ступни были бы более ориентированы медиально (ближе друг к другу), и нагрузка была бы распределена по лучам более равномерно (бороздки, разделяющие более толстые участки стопы), чем у современного человека. Несмотря на эти различия, обнаруженные кости позволяют предположить, что ступни в целом были похожи на ступни современных людей.[69] В 2008 году палеоантропологи Ян Дж. Уоллес, Бриджит Демес, Уильям Л. Юнгерс, Мартин Альверо и Энн Су заявили, что, по их мнению, окаменелости Дманиси были слишком фрагментарными, чтобы сделать вывод о положении ступней (как медиальном) с такой большой уверенностью. полагая, что больше окаменелостей, особенно таз и дополнительные кости стопы.[70]
Палеоэкология



Все окаменелости, обнаруженные в Дманиси, относятся к относительно короткому временному интервалу и представляют собой «снимок во времени».[4] За единственным исключением черепа 5 и его нижней челюсти (которые несколько более ранние по возрасту), все окаменелости гомининов современны, и все окаменелости (включая череп 5), вероятно, откладываются в течение временного интервала, возможно всего 10– 100 тысяч лет.[17]
В плейстоцене городище Дманиси должно было располагаться рядом с берегом озера, образовавшимся в результате перекрытия рек Машавера и Пиназаури потоком лавы.[2] Окружающая среда была бы умеренной, относительно влажной и покрытой лесами; с лесными массивами и галерейными лесами, открытыми лугами, кустарниками, древесными саваннами и скалистыми местностями с кустарниковой растительностью. Окружающая среда, которая также пережила бы холодные зимы, была бы совсем не похожа на сухую и жаркую степь Восточная Африка, где жили более ранние (и современные) гоминины.[34] Даже тогда плейстоценовый Дманиси был, вероятно, теплее и суше, чем нынешняя Грузия, возможно, сравним с средиземноморский климат.[71][72]
Хотя большинство сохранившихся окаменелостей животных предполагают преимущественно лесостепную экосистему, некоторые части фаунистического сообщества подчеркивают, что части окружающей среды были полностью степными (как показано страус и пищуха окаменелости) и сплошной лес (как показано на олень окаменелости).[73] Леса, вероятно, покрывали горные возвышенности и земли вдоль русел рек, тогда как равнинные речные долины были покрыты степной растительностью.[71] Поскольку окаменелости оленей особенно распространены (составляют около 80% окаменелостей, обнаруженных в Дманиси), вероятно, что леса были доминирующим типом окружающей среды.[74]

Окаменелости животных, обнаруженные в тех же отложениях, что и останки гомининов, демонстрируют, что плейстоценовый Дманиси был домом для очень разнообразной фауны.[34] в том числе пищухи,[73] ящерицы, хомяки, черепахи, зайцы, шакалы, медведи, ягуары, носороги, лошадиные, жирафы, коровы и лань.[15] Большинство найденных животных Виллафранкский (а Возраст европейских наземных млекопитающих ) представлены млекопитающие и несколько вымерших видов, в том числе Мегантереон мегантереон и Homotherium crenatidens (и то и другое саблезубые кошки ), Panthera gombaszoegensis (европейский ягуар), Ursus etruscus (этрусский медведь), Equus stenonis (зебра Стенон), Stephanorhinus etruscus (этрусский носорог), Pachystruthio dmanisensis (гигантский страус), олени Cervus perrieri и Cervidae ср. Арверноцерос, гиена Pliocrocuta perrieri, грызуны Mimomys tornensis, М. ostramosensis и Ковальския sp., Газелла ср. борбоника (газель), коза-антилопа Soergelia sp., зубр Бизон георгикус и жираф Жирафы ср. Palaeotraginae.[75] Совместное появление множества крупных плотоядных животных; Мегантереон, Гомотерий, Пантера и Плиокрокута, подчеркивает, что окружающая среда должна была быть весьма разнообразной.[73] Деятельность хищников может быть объяснением того факта, что все черепа гомининов были найдены на расстоянии всего нескольких квадратных метров друг от друга.[17]
Большое количество окаменелых семян растений также было обнаружено в Дманиси, в основном из Boraginaceae и свекла растения. Большинство идентифицированных растений являются современными видами, которые являются несъедобными, хотя присутствовали некоторые съедобные растения, такие как Celtis (каркасы) и Эфедра. В сочетании с Celtis семена часто встречаются и на других участках гомининов (особенно Таутавель во Франции и Чжоукоудянь в Китае), возможно, что каркасы (а также, возможно, Эфедра) были съедены гомининами Дманиси. Обилие семян Boraginaceae, которые часто использовались в более поздних местах как признак человеческой деятельности, могло означать, что гоминины уже оказывали влияние на местную флору в это раннее время.[76] Помимо ягод и фруктов, гоминины, вероятно, были способны использовать широкий спектр ресурсов в пищу. Скорее всего, мясо составляло основную часть их рациона, особенно зимой, когда было труднее найти другие источники пищи.[46]
Большинство окаменелостей (включая все окаменелости гомининов) были извлечены из четвертого из пяти слоев на этом участке, причем в верхних (несколько более молодых) слоях сохранились более поздние отложения. Слои 2 и 3 содержат значительно меньше окаменелостей, почти не сохраняют окаменелости плотоядных животных и останков грызунов или рептилий. Хотя отчасти это может быть связано с предвзятость сохранения, это, вероятно, также отражает некоторые палеоэкологические изменения, вероятно совпавшие с аридизацией Восточной Грузии в раннем плейстоцене.[73] Аридизация повлекла за собой значительное сокращение площади лесных массивов и дальнейшее распространение открытой растительности и степной среды.[71]
Культура
Технологии

В Дманиси было обнаружено более 10 000 каменных орудий, и их стратиграфическая и пространственная концентрация предполагает сложную запись нескольких повторных оккупаций на этом месте. Большинство восстановленных инструментов чешуйчатые инструменты, но меньшее количество каменные ядра и чопперы также были восстановлены. Сырье для изготовления этих каменных орудий, вероятно, происходило из рек и обнажений вблизи места окаменелости.[34][77] Наличие ядер, отщепов и кусков в дополнение к готовым инструментам показывает, что все этапы стучать (формирование камня для создания инструментов) происходило в Дманиси. Хотя техника была не очень сложной, качественные камни (такие как вулканический, магматический и осадочные камни а также окремненный туф ) были использованы. Точная используемая техника различалась от камня к камню, в зависимости от формы исходного камня. Похоже, что в процессе не было создано никаких новых углов.[77]
Хлопья и измельчители обычно считаются довольно простыми инструментами, а инструменты, найденные в Дманиси, во многом аналогичны инструментам олдованской традиции, созданным гомининами в Африке, по крайней мере, почти миллион лет назад. В дополнение к инструментам, найденным на этом месте, также было обнаружено много немодифицированных камней, которые, должно быть, возникли где-то в другом месте из-за их минералогического состава (что означает, что они не прибыли туда естественным путем, а были принесены гомининами). Более крупные немодифицированные камни могли использоваться в качестве инструментов для разбивания костей, резки мяса и истирания плоти, тогда как более мелкие камни могли использоваться для других целей, например, для метания.[34]
Большие коллекции Manuports (немодифицированные камни, перемещенные из их естественного контекста), обнаруженные в Дманиси, обычно интерпретируются как запасы камня, созданные гомининами, чтобы избежать повторных посещений мест сбора камней.[77]
Социальное сотрудничество
Маленькие гоминины Дманиси, возможно, использовали агрессивную уборку мусора, бросая небольшие камни, чтобы украсть пищу у местных хищников. Возможно, что это очищение власти проводилось в группах для защиты, и это могло привести к развитию социального сотрудничества, зависящего от родства.[34]
Есть также косвенное свидетельство социального сотрудничества в Черепе 4, которое принадлежит человеку, который к моменту своей смерти потерял все, кроме одного зуба.[34] Пожилой человек мог бы прожить относительно долгое время после потери зубов, на что указывало то, что лунки корней зубов были заполнены костной тканью, что возможно только в том случае, если рассматриваемый человек жив.[18] Без огня для приготовления пищи беззубому человеку было бы трудно выжить в течение нескольких лет в периодически холодной окружающей среде.Хотя возможно, что с помощью молотков он выжил бы сам, поедая мягкие ткани животных, такие как мозг и костный мозг, более убедительной возможностью является то, что о нем могли заботиться другие представители его вида.[34]
использованная литература
- ^ Pontzer et al. 2010 г., п. 492.
- ^ а б c d е Лордкипанидзе 2017, п. 50.
- ^ а б c d е ж г час я j k л м Августи 2018, п. 74.
- ^ а б c d е ж г час я Лордкипанидзе 2017, п. 49.
- ^ а б c d е Векуа и Лордкипанидзе 2010, п. 161.
- ^ Лордкипанидзе и др. 2007 г., п. 305.
- ^ а б Ферринг и др. 2011 г., п. 1.
- ^ а б c d Лордкипанидзе и др. 2013, п. 326.
- ^ Габуния и др. 2000 г., п. 25.
- ^ а б c Райтмайр, Лордкипанидзе и Векуа 2006, п. 140.
- ^ а б Zollikofer et al. 2014 г., п. 360-б.
- ^ а б Шварц, Таттерсолл и Чи 2014, п. 360-а.
- ^ а б Робертс 2018 С. 110–111.
- ^ а б Лордкипанидзе 2017, п. 48.
- ^ а б c d е ж Векуа и Лордкипанидзе 2010, п. 159.
- ^ а б c Векуа и Лордкипанидзе 2010, п. 160.
- ^ а б c Скиннер, Гордон и Коллард, 2006 г., п. 43.
- ^ а б c Векуа и Лордкипанидзе 2010, п. 162.
- ^ Габуния и др. 2000 г., п. 15.
- ^ а б Лордкипанидзе и др. 2007 г., п. 306.
- ^ а б Лордкипанидзе и др. 2007 г., п. 307.
- ^ а б c d Лордкипанидзе и др. 2007 г., п. 308.
- ^ а б Габуния и др. 2000 г..
- ^ а б c d Габуния и др. 2002 г., п. 244.
- ^ а б c d Rightmire et al. 2017 г., п. 54.
- ^ Габуния и Векуа 1995.
- ^ а б Vekua et al. 2002 г., п. 88.
- ^ Vekua et al. 2002 г..
- ^ Vekua et al. 2002 г., п. 86.
- ^ Лордкипанидзе и др. 2006 г., п. 1150.
- ^ Лордкипанидзе и др. 2005 г..
- ^ Лордкипанидзе и др. 2006 г..
- ^ Лордкипанидзе и др. 2005 г., п. 718.
- ^ а б c d е ж г час Лордкипанидзе 2017, п. 51.
- ^ Лордкипанидзе и др. 2013.
- ^ Габуния и др. 2002 г..
- ^ Габуния и Векуа 1995, п. 509.
- ^ Габуния и Векуа 1995, п. 510.
- ^ Bräuer & Schultz 1996, п. 478.
- ^ Росас и Кастро 1998, п. 145.
- ^ Габуния и др. 2000 г., п. 1020.
- ^ Габуния и др. 2000 г., п. 1021.
- ^ Габуния, Векуа и Лордкипанидзе 2000, п. 790.
- ^ Габуния и др. 2002 г., п. 245.
- ^ Ли 2005, п. 263.
- ^ а б Лордкипанидзе и др. 2006 г., п. 1156.
- ^ Райтмайр, Лордкипанидзе и Векуа 2006, п. 130.
- ^ Райтмайр, Лордкипанидзе и Векуа 2006.
- ^ Скиннер, Гордон и Коллард, 2006 г., п. 45.
- ^ Райтмайр, Ван Арсдейл и Лордкипанидзе, 2008 г., п. 907.
- ^ Райтмайр, Ван Арсдейл и Лордкипанидзе, 2008 г., п. 908.
- ^ Мартинон-Торрес и др. 2008 г., п. 269.
- ^ Macaluso 2010, п. 38.
- ^ Лордкипанидзе и др. 2013, п. 329.
- ^ Лордкипанидзе и др. 2013, п. 330.
- ^ Rightmire et al. 2017 г., п. 62.
- ^ Rightmire et al. 2017 г., п. 74.
- ^ Rightmire et al. 2017 г., п. 75.
- ^ Rightmire et al. 2017 г., п. 76.
- ^ Габуния и др. 2000 г., п. 1019.
- ^ а б Гарсия и др. 2010 г., п. 449.
- ^ Zhu et al. 2018 г., п. 608.
- ^ Габуния, Векуа и Лордкипанидзе 2000, п. 798.
- ^ Rightmire et al. 2017 г., п. 70.
- ^ а б c Лордкипанидзе и др. 2013, п. 327.
- ^ Лордкипанидзе 2017, п. 52.
- ^ Rightmire et al. 2017 г., п. 61.
- ^ Pontzer et al. 2010 г., п. 503.
- ^ а б Лордкипанидзе и др. 2007 г., п. 309.
- ^ Wallace et al. 2008 г., п. 377.
- ^ а б c Габуния и др. 2000 г., п. 24.
- ^ Габуния, Векуа и Лордкипанидзе 2000, п. 793.
- ^ а б c d Габуния и др. 2000 г., п. 22.
- ^ Габуния, Векуа и Лордкипанидзе 2000, п. 795.
- ^ Габуния и др. 2000 г. С. 19–21.
- ^ Габуния, Векуа и Лордкипанидзе 2000, п. 797.
- ^ а б c Мгеладзе и др. 2011 г., п. 593.
Цитированная библиография
- Агусти, Хорди (2018). «Эволюция рода Homo: новые загадки и перспективы». Журнал научных исследований Mètode. 8: 71–77.
- Бройер, Гюнтер; Шульц, Майкл (1996). «Морфологическое сходство нижней челюсти плио-плейстоцена из Дманиси, Грузия». Журнал эволюции человека. 30 (5): 445–481. Дои:10.1006 / jhev.1996.0037.
- Габуния, Лев; Векуа, Абесалом (1995). "Плио-плейстоценовый гоминид из Дманиси, Восточная Грузия, Кавказ". Природа. 373 (6514): 509–512. Bibcode:1995Натура 373..509G. Дои:10.1038 / 373509a0. PMID 7845461. S2CID 4242943.
- Габуния, Лев; и другие. (2000). «Самые ранние плейстоценовые черепные останки гоминидов из Дманиси, Республика Грузия: таксономия, геологические условия и возраст». Наука. 288 (5468): 1019–1025. Bibcode:2000Sci ... 288.1019G. Дои:10.1126 / science.288.5468.1019. PMID 10807567.
- Габуния, Лев; и другие. (2000). "Текущие исследования местонахождения гоминидов в Дманиси" (PDF). ERAUL. 92: 13–27.
- Габуния, Лев; Векуа, Абесалом; Лордкипанидзе, Давид (2000). «Экологические контексты ранней оккупации Грузии (Закавказья)». Журнал эволюции человека. 38 (6): 785–802. Дои:10.1006 / jhev.1999.0383. PMID 10835262.
- Габуния, Лев; де Ламли, Мария-Антуанетта; Векуа, Абесалом; Лордкипанидзе, Давид; де Ламли, Генри (2002). «Обнаружение нового гоминида в Дманиси (Закавказье, Грузия)». Comptes Rendus Palevol. 1 (4): 243–253.
- Гарсия, Тристан; Феро, Жильбер; Фальгер, Кристоф; де Ламли, Генри; Перрену, Кристиан; Лордкипанидзе, Давид (2010). «Самые ранние человеческие останки в Евразии: Новое 40Ar /39Датирование уровней гоминидности Дманиси, Грузия ". Четвертичная геохронология. 5 (4): 443–451. Дои:10.1016 / j.quageo.2009.09.012.
- Ферринг, Рид; Омс, Ориэл; Агусти, Хорди; Берна, Франческо; Ниорадзе, Медея; Шелия, Теона; Таппен, Марта; Векуа, Абесалом; Жвания, Давид; Лордкипанидзе, Давид (2011). «Самые ранние занятия людей в Дманиси (Грузинский Кавказ) датируются 1.85–1.78 млн лет назад». Труды Национальной академии наук Соединенных Штатов Америки. 108 (26): 10432–6. Дои:10.1073 / pnas.1106638108. ЧВК 3127884. PMID 21646521.
- Лебатар, Анна-Элизабет; и другие. (2013). "Знакомства человек прямоходящий с травертином из Коджабаша (Денизли, Турция) минимум 1,1 млн лет назад. Письма по науке о Земле и планетах. 390: 8–18. Дои:10.1016 / j.epsl.2013.12.031.
- Ли, Санг-Хи (2005). «Краткое сообщение: вариация емкости черепа образца Дманиси слишком велика, чтобы происходить от одного вида?» (PDF). Американский журнал физической антропологии. 127 (3): 263–266. Дои:10.1002 / ajpa.20105. PMID 15503340.
- Лордкипанидзе, Давид; и другие. (2005). «Самый ранний череп беззубого гоминина». Природа. 434 (7034): 717–718. Дои:10.1038 / 434717b. PMID 15815618. S2CID 52800194.
- Лордкипанидзе, Давид; и другие. (2006). «Четвертый череп гоминина из Дманиси, Грузия». Анатомическая запись, часть А. 288A (11): 1146–1157. Дои:10.1002 / ar.a.20379. PMID 17031841.
- Лордкипанидзе, Давид; и другие. (2007). "Посткраниальные свидетельства раннего Гомо из Дманиси, Грузия ". Природа. 449 (7160): 305–310. Bibcode:2007Натура.449..305л. Дои:10.1038 / природа06134. PMID 17882214.
- Лордкипанидзе, Давид; Понсе де Леон, Марсия С .; Маргвелашвили, Энн; Рак, Йоэль; Райтмайр, Дж. Филип; Векуа, Абесалом; Цолликофер, Кристоф П. Э. (2013). "Полный череп из Дманиси, Грузия, и эволюционная биология раннего Гомо". Наука. 342 (6156): 326–331. Bibcode:2013Наука ... 342..326Л. Дои:10.1126 / science.1238484. PMID 24136960.
- Лордкипанидзе, Давид (2017). «История раннего Гомо". В Тибайренке, Мишель; Айяла, Франсиско Дж. (Ред.). О природе человека: биология, психология, этика, политика и религия. Академическая пресса. С. 45–54. ISBN 978-0-12-420190-3.
- Макалузо младший, П. Джеймс (2010). «Вариация в останках зубов из Дманиси, Грузия». Антропологическая наука. 118 (1): 31–40. Дои:10.1537 / ase.090501.
- Мартинон-Торрес, Мария; Бермудес де Кастро, Хосе Мария; Гомес-Роблес, Аида; Маргвелашвили, Энн; Прадо, Лейр; Лордкипанидзе, Давид; Векуа, Абесалом (2008). «Стоматологические останки из Дманиси (Республика Грузия): морфологический анализ и сравнительное исследование». Журнал эволюции человека. 55 (2): 249–273. Дои:10.1016 / j.jhevol.2007.12.008. PMID 18486183.
- Мгеладзе, Ана; Лордкипанидзе, Давид; Монсель, Мари-Элен; Despriee, Джеки; Чагелишвили, Русудан; Ниорадзе, Медея; Ниорадзе, Георгий (2011). «Занятия гомининов на территории Дманиси, Грузия, Южный Кавказ: сырье и техническое поведение первых гомининов Европы». Журнал эволюции человека. 60 (5): 571–596. Дои:10.1016 / j.jhevol.2010.10.008. PMID 21277002.
- Понцер, Герман; Ролиан, Кэмпбелл; Райтмайр, Дж. Филип; Джашашвили, Чай; Понсе де Леон, Марсия С .; Лордкипанидзе, Давид; Цолликофер, Кристоф П. Э. (2010). «Опорно-двигательная анатомия и биомеханика гоминидов из Дманиси». Журнал эволюции человека. 58 (6): 492–504. Дои:10.1016 / j.jhevol.2010.03.006. PMID 20447679.
- Райтмайр, Дж. Филип; Лордкипанидзе, Давид; Векуа, Абесалом (2006). «Анатомические описания, сравнительные исследования и эволюционное значение черепов гомининов из Дманиси, Республика Грузия». Журнал эволюции человека. 50 (2): 115–141. Дои:10.1016 / j.jhevol.2005.07.009. PMID 16271745.
- Райтмайр, Дж. Филип; Ван Арсдейл, Адам П .; Лордкипанидзе, Давид (2008). «Вариация нижних челюстей из Дманиси, Грузия». Журнал эволюции человека. 54 (6): 904–908. Дои:10.1016 / j.jhevol.2008.02.003. PMID 18394678.
- Райтмайр, Дж. Филип; Понсе де Леон, Марсия С .; Лордкипанидзе, Давид; Маргвелашвили, Энн; Цолликофер, Кристоф П. Э. (2017). «Череп 5 из Дманиси: описательная анатомия, сравнительные исследования и эволюционное значение». Журнал эволюции человека. 104: 50–79. Дои:10.1016 / j.jhevol.2017.01.005. PMID 28317556.
- Робертс, Алиса (2018). Эволюция: история человечества (Пересмотренная ред.). Dorling Kindersley Ltd. ISBN 978-0-2413-0431-0.
- Росас, Антонио; Де Кастро, Хосе Мария Бермудес (1998). «О таксономическом сходстве нижней челюсти Дманиси (Грузия)». Американский журнал физической антропологии. 107 (2): 145–162. Дои:10.1002 / (SICI) 1096-8644 (199810) 107: 2 <145 :: AID-AJPA2> 3.0.CO; 2-U. PMID 9786330.
- Шварц, Джеффри Х .; Таттерсолл, Ян; Чи, Чжан (2014). "Комментарий к" Полный череп из Дманиси, Грузия, и эволюционная биология раннего Гомо"". Наука. 344 (6182): 360 – а. Bibcode:2014Наука ... 344..360С. Дои:10.1126 / science.1250056. PMID 24763572. S2CID 36578190.
- Скиннер, Мэтью М .; Гордон, Адам Д .; Коллард, Николь Дж. (2006). «Изменение размера и формы нижней челюсти у гомининов в Дманиси, Республика Грузия». Журнал эволюции человека. 51 (1): 36–49. Дои:10.1016 / j.jhevol.2006.01.006. PMID 16563468.
- Векуа, Абесалом; и другие. (2002). "Новый череп раннего Гомо из Дманиси, Грузия ". Наука. 297 (5578): 85–89. Bibcode:2002Наука ... 297 ... 85В. Дои:10.1126 / science.1072953. PMID 12098694.
- Векуа, Абесалом; Лордкипанидзе, Давид (2010). «Дманиси (Грузия) - место открытия старейших гоминидов в Евразии» (PDF). Вестник Национальной академии наук Грузии. 4 (2): 158–164.
- Уоллес, Ян Дж .; Демес, Бриджит; Юнгерс, Уильям Л .; Альверо, Мартин; Су, Энн (2008). "Двуногие гоминины Дманиси: голубиногороды в начале Гомо?". Американский журнал физической антропологии. 136 (4): 375–378. Дои:10.1002 / ajpa.20827. PMID 18350586.
- Чжу, Чжаоюй; Деннелл, Робин; Хуанг, Вэйвэнь; Ву, Йи; Цю, Шифань; Ян, Шися; Рао, Чжиго; Хоу, Ямэй; Се, Цзюбинь; Хан, Цзянвэй; Оуян, Тинпин (2018). «Гомининское заселение китайского Лессового плато примерно 2,1 миллиона лет назад». Природа. 559 (7715): 608–612. Bibcode:2018Натура.559..608Z. Дои:10.1038 / s41586-018-0299-4. PMID 29995848. S2CID 49670311.
- Цолликофер, Кристоф П. Э .; Понсе де Леон, Marca S .; Маргвелашвили, Энн; Райтмайр, Дж. Филип; Лордкипанидзе, Давид (2014). "Ответ на комментарий" Полный череп из Дманиси, Грузия, и эволюционная биология раннего Гомо"". Наука. 344 (6182): 360 – b. Bibcode:2014Научный ... 344..360Z. Дои:10.1126 / science.1250081. PMID 24763573. S2CID 206554612.
внешние ссылки
- Dmanisi.ge - веб-сайт, посвященный исследованиям гомининов Дманиси и месту ископаемых останков Дманиси
- Homo georgicus at Milne Publishing - История нашего племени: Гоминини
- Человек прямоходящий georgicus at Origins - Изучение летописи окаменелостей - Фонд Брэдшоу
| Таксономия (Гоминины ) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Предки |
| ||||||||||||||||||||||||||
| Модели |
| ||||||||||||||||||||||||||
| Сроки | |||||||||||||||||||||||||||
| Другие |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||