Недавнее африканское происхождение современного человека - Recent African origin of modern humans
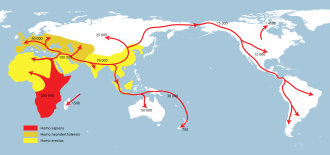
человек прямоходящий наибольшая степень (желтый),
Homo neanderthalensis наибольшая степень (охра) и
Homo sapiens (красный).


В палеоантропология, то недавнее африканское происхождение современного человека, также называемый "Из Африки" теория (OOA), недавняя гипотеза единственного происхождения (RSOH), гипотеза замены, или же недавняя модель африканского происхождения (РАО), является доминирующим[1][2][3] модель географического происхождения и ранняя миграция из анатомически современные люди (Homo sapiens ). Это следует за ранние экспансии гомининов из Африки, выполненный человек прямоходящий а потом Homo neanderthalensis.
Модель предлагает «единое происхождение» Homo sapiens в таксономическом смысле, исключаяпараллельная эволюция рассматриваемых черт анатомически современный в других регионах,[4] но не исключает множественных примесь между Х. сапиенс и архаичные люди в Европе и Азии.[примечание 1][5][6] Х. сапиенс скорее всего, разработан в Африканский рог между 300 000 и 200 000 лет назад.[7][8] Модель «недавнего африканского происхождения» предполагает, что все современные неафриканские популяции в значительной степени происходят от популяций Х. сапиенс который покинул Африку после этого.
Было по крайней мере несколько случаев расселения современных людей "из Африки", возможно, начавшееся уже 270 000 лет назад, в том числе 215 000 лет назад по крайней мере до Греция,[9][10][11] и, конечно же, через северную Африку около 130 000–115 000 лет назад.[12][13][14][15][16][17]Эти ранние волны, по-видимому, в основном вымерли или отступили 80 000 лет назад.[18]
Наиболее значительная «недавняя» волна произошла около 70 000–50 000 лет назад,[7][8][19][20][21] через так называемый "Южный маршрут ", быстро распространяясь вдоль побережья Азии и достигая Австралия примерно 65 000–50 000 лет назад,[22][23][заметка 2] (хотя некоторые исследователи ставят под сомнение более ранние австралийские даты и считают, что люди прибыли сюда не ранее 50 000 лет назад,[24][25] в то время как другие предположили, что эти первые поселенцы Австралии могут представлять более старую волну до более значительной миграции из Африки и, следовательно, не обязательно являются предками более поздних жителей региона.[21]), в то время как Европа была заселена ранним ответвлением, заселившим Ближний Восток и Европу менее 55000 лет назад.[26][27][28]
В 2010-х учится в популяционная генетика обнаруженные доказательства скрещивание это произошло между Х. сапиенс и архаичные люди в Евразии, Океании и Африке,[29][30][31] указывает на то, что современные группы населения, хотя в основном произошли от ранних Х. сапиенс, в меньшей степени также произошли от региональных вариантов архаичных людей.
Предлагаемые волны
- Видеть Ранние экспансии гомининов из Африки для архаичных людей (H. erectus, Х. heidelbergensis, Неандертальцы, денисовцы).

«Недавнее африканское происхождение» или Из Африки II, относится к миграции анатомически современные люди (Homo sapiens ) из Африки после их появления в c. 300 000–200 000 лет назад, в отличие от "Из Африки я ", который относится к миграции архаичных людей из Африки в Евразию примерно 1,8–0,5 миллиона лет назад. Омо-Кибиш I (Омо I) из южной Эфиопии - самый старый из известных в настоящее время анатомически современных скелетов Homo sapiens (196 ± 5 тыс. лет назад).[33]
С начала 21 века картина «недавних миграций единственного происхождения» стала значительно более сложной не только из-за открытия примеси современной архаики, но и из-за растущего числа свидетельств того, что «недавние выходящие из-за границы» Африканская миграция происходила в виде нескольких волн, распространявшихся в течение длительного периода времени. По состоянию на 2010 год существовало два основных принятых пути расселения для миграции за пределы Африки ранних анатомически современных людей: через «Северный путь» (через Долина Нила и Синай) и "Южный путь" через Баб-эль-Мандаб пролив.[34]
- Posth et al. (2017) предполагают, что ранние Homo sapiens, или «другой вид в Африке, тесно связанный с нами», возможно, впервые мигрировал из Африки около 270 000 лет назад.[35]
- Находки на Пещера Мислия, которые включают частичную челюстную кость с восемью зубами, датируются примерно 185 000 лет назад. Слои, датируемые периодом от 250 000 до 140 000 лет назад в той же пещере, содержали инструменты Леваллуа типа, который мог бы поставить дату первой миграции еще раньше, если бы инструменты могли быть связаны с современными находками человеческих челюстей.[36][37]
- Распространение на восток от Северо-Восточной Африки до Аравии 150 000–130 000 лет назад на основании находок в Джебель Фая датируется 127000 лет назад (обнаружено в 2011 году).[12][13] Возможно, с этой волной связаны находки из Чжирендун пещера, Южный Китай, датируемая более 100 000 лет назад.[34] Другое свидетельство присутствия современного человека в Китае датируется 80 000 лет назад.[18]
- Наиболее значительное распространение произошло около 50–70 000 лет назад через так называемые Южный маршрут, либо до[38] или после[27][28] в Событие Тоба, что произошло между 69 000 и 77 000 лет назад.[38] Это расселение следовало по южному побережью Азии и достигло Австралии около 65-50 тысяч лет назад, или, согласно некоторым исследованиям, самое раннее 50 тысяч лет назад.[24][25] Западная Азия была «заселена» другим происхождением этой волны около 50 000 лет назад, а Европа была заселена из Западной Азии около 43 000 лет назад.[34]
- Уэллс (2003) описывает дополнительную волну миграции после южного прибрежного маршрута, а именно миграцию с севера в Европу около 45000 лет назад.[заметка 3] Однако такая возможность исключается Macaulay et al. (2005) и Posth et al. (2016), которые выступают за единственное прибрежное рассредоточение с ранним ответвлением в Европу.
Разгон по Северному маршруту

Начиная с 135 000 лет назад в тропической Африке мегапроявления который изгнал людей с суши к морским берегам и заставил их перейти на другие континенты.[39][примечание 4]
Современные люди пересекли пролив Баб-эль-Мандеб на юге красное море, и двинулся вдоль зеленых берегов вокруг Аравии, а оттуда в остальную часть Евразии. Окаменелости ранних Homo sapiens были найдены в Кафзех пещера в Израиле и датируется от 80 000 до 100 000 лет назад. Похоже, что эти люди либо вымерли, либо отступили обратно в Африку 70 000–80 000 лет назад, возможно, их заменили движущиеся на юг неандертальцы, бежавшие из более холодных регионов Европы ледникового периода.[40] Хуа Лю и другие. проанализировали аутосомные микросателлитные маркеры, датируемые примерно 56 000 лет назад. Они интерпретируют палеонтологический ископаемое как изолированное раннее ответвление, вернувшееся в Африку.[41]
Открытие каменных орудий труда в Объединенные Арабские Эмираты в 2011 году на Сайт Фая-1 в Млейха, Шарджа, указывает на присутствие современного человека не менее 125000 лет назад,[12] что привело к возрождению «давно забытого» маршрута в Северной Африке.[13][42][14][15]
В Оман, сайт был обнаружен Бьен Ховен в 2011 году, содержащий более 100 фрагментов поверхности каменных орудий, принадлежащих к позднему нубийскому комплексу, ранее известному только из археологические раскопки в Судан. По двум оценкам возраста с использованием оптически стимулированной люминесценции, возраст Аравийско-Нубийского комплекса составляет примерно 106 000 лет. Это свидетельствует о явном Каменный век технокомплекс на юге Аравии, около ранней части Морской изотоп, стадия 5.[43]
По словам Кулвильма и его соавторов, Неандертальцы внесенный генетически до современных людей, которые тогда жили за пределами Африки около 100000 лет назад: люди, которые уже отделились от других современных людей около 200000 лет назад, и эта ранняя волна современных людей за пределами Африки также генетически внесла свой вклад в развитие алтайских неандертальцев.[44] Они обнаружили, что «предки неандертальцев из Горный Алтай и ранние современные люди встретились и скрестились, возможно, на Ближнем Востоке, на много тысяч лет раньше, чем считалось ранее ".[44] По словам соавтора Илана Гронау, «это фактически дополняет археологические свидетельства присутствия ранних современных людей из Африки около 100 000 лет назад и ранее, предоставляя первые генетические свидетельства существования таких популяций».[44] Похожий генетическая примесь события были отмечены в других регионах также.[45]
В Китае Люцзян человек (Китайский : 柳江 人) является одним из первых современных людей, обнаруженных в Восточная Азия.[46]Дата, которую чаще всего приписывают останкам, - 67000 лет назад.[47] Высокие показатели изменчивости, полученные с помощью различных методов датирования, проведенных разными исследователями, позволяют отнести наиболее широко принятый диапазон дат как минимум к 67 000 лет назад, но не исключают датировки до 159 000 лет назад.[47] Лю, Мартинон-Торрес и др. (2015) утверждают, что современные человеческие зубы были найдены в Китае не менее 80 000 лет назад.[48]
Разгон Южного маршрута
Прибрежный маршрут

Примерно 50-70 000 лет назад часть носителей митохондриальной гаплогруппы L3 перекочевал из Восточная Африка в Ближний Восток. Было подсчитано, что из популяции в Африке от 2000 до 5000 человек только небольшая группа, возможно, всего от 150 до 1000 человек, пересекла Красное море.[49][50] Группа, пересекшая Красное море, двигалась по прибрежному маршруту вокруг Аравия и Персидское плато в Индию, которая, по-видимому, была первым крупным пунктом урегулирования.[51] Уэллс (2003) выступал за маршрут вдоль южного побережья Азии примерно через 250 километров (155 миль)[сомнительный ], достигнув Австралии около 50 000 лет назад.
Сегодня в Баб-эль-Мандебский пролив, то красное море имеет ширину около 20 километров (12 миль), но 50 000 лет назад уровень моря был на 70 м (230 футов) ниже (из-за оледенения), а вода была намного уже. Хотя проливы никогда не закрывались полностью, они были достаточно узкими, чтобы их можно было пересечь на простых плотах, и между ними могли быть острова.[52][34] Ракушка кучи 125000 лет были найдены в Эритрея,[53] указывает на то, что в рацион древних людей входили морепродукты, полученные пляж.
Датировка Южного Расселения является предметом споров.[38] Это могло произойти либо до, либо после Тоба, катастрофического извержения вулкана, которое произошло между 69000 и 77000 лет назад на месте современного Озеро Тоба. Каменные орудия, обнаруженные под слоями пепла в Индии, могут указывать на расселение до Тоба, но источник этих орудий оспаривается.[38] Признаком пост-Тоба является гаплогруппа L3, которая возникла до расселения людей из Африки и может быть датирована 60 000–70 000 лет назад, «предполагая, что человечество покинуло Африку через несколько тысяч лет после Тоба».[38] Некоторые исследования, показывающие более медленные, чем ожидалось, генетические мутации в ДНК человека, были опубликованы в 2012 году, что указывает на пересмотренную датировку миграции между 90 000 и 130 000 лет назад.[54] Некоторые более поздние исследования предполагают миграцию из Африки около 50 000–65 000 лет назад предков современных неафриканских популяций, что аналогично большинству предыдущих оценок.[21][55][56]
Западная Азия
Окаменелость современного человека, датированная 54700 лет назад, была найдена в Пещера Манот в Израиле по имени Манот 1,[57] хотя датировка была подвергнута сомнению Groucutt et al. (2015).
Южная Азия и Австралия
Считается, что Австралия была заселена примерно 65 000–50 000 лет назад. По состоянию на 2017 год самым ранним свидетельствам существования людей в Австралии не менее 65000 лет.[22][23] в то время как Макчесни заявил, что
... генетические данные свидетельствуют о том, что небольшая группа с маркером M168 мигрировала из Африки вдоль побережья Аравийского полуострова и Индии через Индонезию и достигла Австралии очень рано, между 60 000 и 50 000 лет назад. Эта очень ранняя миграция в Австралию также подтверждается Rasmussen et al. (2011).[26]
Окаменелости из Озеро Мунго, Австралия, были датированы примерно 42 000 лет назад.[58][59] Другие окаменелости с участка под названием Маджедбебе были датированы по крайней мере 65000 лет назад.,[23] хотя некоторые исследователи сомневаются в этой ранней оценке и датируют окаменелости Мадджедбебе примерно 50 000 лет назад самым старым.[24][25]
Восточная Азия
Тяньюань человек из Китай имеет вероятный диапазон дат между 38000 и 42000 лет назад, в то время как Люцзян человек из того же региона имеет вероятный диапазон дат от 67 000 до 159 000 лет назад. Согласно тестам ДНК 2013 года, человек Тяньюань связан "со многими современными людьми. Азиаты и Коренные американцы ".[60][61][62][63][64] Тяньюань похож на морфология к Человек Минатогава, современные люди датируются периодом от 17000 до 19000 лет назад и найдены на Окинава Остров, Япония.[65][66]
Европа
В соответствии с Macaulay et al. (2005), раннее ответвление от южного расселения с гаплогруппой N, следовало за Нилом из Восточной Африки, направляясь на север и пересекая Азия сквозь Синай. Затем эта группа разветвлялась: одни двинулись в Европу, а другие - на восток, в Азию.[27] Эта гипотеза подтверждается относительно поздней датой прибытия современных людей в Европу, а также археологическими данными и данными ДНК.[27] На основе анализа 55 митохондриальных геномов человека (мтДНК) охотников-собирателей, Posth et al. (2016) приводят доводы в пользу «быстрого однократного расселения всех неафриканцев менее 55 000 лет назад».
Генетическая реконструкция
Митохондриальные гаплогруппы
В Африке

Первая линия, от которой ответвляется Митохондриальная Ева был L0. Эта гаплогруппа широко распространена среди Сан юга Африки и Сандаве Восточной Африки. Он также встречается среди Мбути люди.[67][68] Эти группы разделились на раннем этапе истории человечества и с тех пор оставались относительно генетически изолированными. Гаплогруппы L1, L2 и L3 являются потомками L1 – L6 и в основном обитают в Африке. Макрогаплогруппы M и N, которые являются потомками остального мира за пределами Африки, происходят от L3. L3 около 70 000 лет, а гаплогруппам M и N около 65-55 000 лет.[69][56] Взаимосвязь между такими генетическими деревьями и демографической историей все еще обсуждается применительно к расселению.[70]
Из всех родословных, присутствующих в Африке, только потомки женского пола одной линии, мтДНК гаплогруппа L3, находятся за пределами Африки. Если бы было несколько миграций, можно было бы ожидать, что будут найдены потомки более чем одной линии. Потомки L3 женского пола, M и N линии гаплогруппы встречаются в Африке с очень низкой частотой (хотя гаплогруппа M1 популяции очень древние и разнообразны по север и Северо-Восточная Африка ) и, похоже, более поздние.[нужна цитата ] Возможное объяснение состоит в том, что эти мутации произошли в Восточной Африке незадолго до массового исхода и впоследствии стали доминирующими гаплогруппами посредством эффект основателя. С другой стороны, мутации могли возникнуть вскоре после этого.
Южный маршрут и гаплогруппы M и N
Результаты мтДНК, собранной у аборигенов Малайзии, назвали Оранг Асли указывают на то, что гапологруппы M и N имеют общие характеристики с оригинальными африканскими группами примерно 85000 лет назад и имеют общие характеристики с субгаплогруппами, обнаруженными в прибрежных регионах Юго-Восточной Азии, таких как Австралазия, Индийский субконтинент и во всей континентальной Азии, которые рассредоточились и отделился от своего африканского прародителя примерно 65 000 лет назад. Это распространение на южном побережье могло произойти до распространения через Левант примерно 45 000 лет назад.[27] Эта гипотеза пытается объяснить, почему гаплогруппа N преобладает в Европе и почему гаплогруппа M отсутствует в Европе. Предполагается, что свидетельства прибрежной миграции были уничтожены повышением уровня моря во время Голоцен эпоха.[71] С другой стороны, небольшая европейская популяция основателей, которая сначала выражала гаплогруппу M и N, могла потерять гаплогруппу M из-за случайной генетический дрейф в результате горлышко бутылки (т.е. эффект основателя ).
Группа, пересекшая Красное море, двигалась по прибрежному маршруту вокруг Аравия и Персия до достижения Индии.[51] Гаплогруппа M часто встречается в южных прибрежных районах Пакистан и Индия, и наибольшее разнообразие в Индии, что указывает на то, что именно здесь, возможно, произошла мутация.[51] Шестьдесят процентов населения Индии принадлежат к Гаплогруппа M. Коренные жители Андаманские острова также принадлежат к линии М. Считается, что андаманцы являются ответвлениями некоторых из первых жителей Азии из-за их долгой изоляции от материка. Они свидетельствуют о прибрежном маршруте первых поселенцев, который простирается от Индии до Таиланд и Индонезия до востока Новая Гвинея. Поскольку M встречается в высоких частотах у горцев из Новой Гвинеи, а андаманцы и жители Новой Гвинеи имеют темную кожу и Афро-текстурированные волосы, некоторые ученые думают, что все они являются частью той же волны мигрантов, которые пересекли Красное море ~ 60 000 лет назад в Великая прибрежная миграция. Доля гаплогруппы M увеличивается к востоку от Аравия в Индию; в восточной Индии M превосходит N в соотношении 3: 1. Пересекая Юго-Восточную Азию, гаплогруппа N (в основном в форме производных от ее субклада R) снова появляется как преобладающая линия.[нужна цитата ] M преобладает в Восточной Азии, но среди Коренные австралийцы, N - более распространенная линия.[нужна цитата ] Такое случайное распространение гаплогруппы N из Европы в Австралию можно объяснить тем, что эффекты основателя и узкие места населения.[72]
Аутосомная ДНК
Исследование населения Африки, Европы и Азии в 2002 году выявило большее генетическое разнообразие среди африканцев, чем среди евразийцев, и что генетическое разнообразие среди евразийцев в значительной степени является подмножеством такового среди африканцев, что подтверждает модель «за пределами Африки».[73] Большое исследование Купа и другие. (2009) нашли доказательства естественный отбор в аутосомный ДНК за пределами Африки. В исследовании выделяются неафриканские зачистки (особенно KITLG варианты, связанные с цвет кожи ), Западно-Евразийские стреловидности (SLC24A5 ) и восточноазиатские стреловидности (MC1R, актуально для цвета кожи). Основываясь на этих доказательствах, исследование пришло к выводу, что человеческие популяции столкнулись с новым селективным давлением по мере их распространения за пределы Африки.[74] MC1R и его связь с цветом кожи уже обсуждалась Лю, Хардинг и др. (2000), п. 135. Согласно этому исследованию, жители Папуа-Новой Гвинеи продолжали подвергаться селекции по темному цвету кожи, так что, хотя эти группы отличаются от африканцев в других местах, аллель темного цвета кожи, разделяемый современными африканцами, андаманцами и новогвинейцами, является архаизмом. . Endicott et al. (2003) предложить конвергентная эволюция. Исследование 2014 года, проведенное Гурдасани и соавт. указывает на то, что более высокое генетическое разнообразие в Африке было дополнительно увеличено в некоторых регионах в результате относительно недавних миграций из Евразии, затронувших некоторые части Африки.[75]
Патогенная ДНК
Еще один многообещающий путь к реконструкции генетической генеалогии человека - через JC вирус (JCV), тип человека полиомавирус который переносится 70–90 процентами людей и обычно передается вертикально от родителей к потомству, что свидетельствует о совместном расхождении с человеческими популяциями. По этой причине JCV использовался в качестве генетического маркера эволюции и миграции человека.[76] Этот метод не представляется надежным для миграции из Африки, в отличие от генетики человека, штаммы JCV, связанные с африканскими популяциями, не являются базальными. Из этого Shackelton et al. (2006) пришли к выводу, что либо базальный африканский штамм JCV вымер, либо первоначальная инфекция JCV возникла после миграции из Африки.
Смесь архаичного и современного человека
Доказательства для архаичный человек виды (произошли от Homo heidelbergensis ) имея скрещенный с современными людьми за пределами Африки, была обнаружена в 2010-х годах. Это касается прежде всего Неандерталец примесь во всех современных популяциях, кроме Африканцы к югу от Сахары но также были представлены доказательства Денисова гоминин примесь в Австралазия (т.е. в Меланезийцы, Австралийские аборигены и немного Негритос ).[77]
Скорость поступления примеси неандертальцев в европейское и азиатское население по состоянию на 2017 год оценивается примерно в 2–3%.[78]
Архаическая примесь в некоторых группах охотников-собирателей африканского населения к югу от Сахары (Биака Пигмеи и Сан ), полученный из архаичных гомининов, которые отделились от современной человеческой линии примерно 700 000 лет назад, был обнаружен в 2011 году. Уровень примеси оценивался примерно в 2%.[31]Примесь архаичных гомининов еще более раннего времени расхождения, оцененного от 1,2 до 1,3 миллиона лет назад, была обнаружена в Пигмеи, Хадза и пять Сандаве в 2012.[79][30]Из анализа Муцин 7, сильно дивергентный гаплотип, который имеет предполагаемое время слияния с другими вариантами около 4,5 миллионов лет назад и специфичен для африканских популяций, предположительно был получен в результате скрещивания между африканскими современными и архаичными людьми.[80]
Каменные инструменты
Помимо генетического анализа, Petraglia и другие. также исследует мелкие каменные орудия труда (микролитический материалы) с Индийского субконтинента и объясняет рост населения на основе реконструкции палеосреды. Он предположил, что каменные орудия в Южной Азии могут быть датированы 35 тыс. Лет назад, а на новую технологию могут повлиять изменения окружающей среды и давление населения.[81]
История теории
Классическая палеоантропология

Кладистические отношения людей с африканцами обезьяны было предложено Чарльз Дарвин после изучения поведения африканцев обезьяны, один из которых был показан на Лондонский зоопарк.[82] Анатом Томас Хаксли также поддержал эту гипотезу и предположил, что африканские обезьяны имеют тесные эволюционные отношения с людьми.[83] Против этих взглядов выступил немецкий биолог. Эрнст Геккель, который был сторонником Теория из Азии. Геккель утверждал, что люди более тесно связаны с приматами Юго-Восточной Азии, и отверг африканскую гипотезу Дарвина.[84][85]
в Происхождение человека Дарвин предположил, что люди произошли от обезьян, у которых все еще был маленький мозг, но они ходили прямо, освобождая руки для целей, способствующих развитию интеллекта; он думал, что такие обезьяны были африканскими:
В каждом великом регионе мира живые млекопитающие тесно связаны с вымершими видами того же региона. Следовательно, вероятно, что в Африке раньше жили вымершие обезьяны, тесно связанные с горилла и шимпанзе; и поскольку эти два вида теперь являются ближайшими союзниками человека, несколько более вероятно, что наши ранние предки жили на африканском континенте, чем где-либо еще. Но рассуждать на эту тему бесполезно, ведь обезьяна размером почти с человека, а именно Дриопитек Ларте, который был тесно связан с антропоморфными Гилобаты, существовавшие в Европе в Верхний миоцен период; и с тех пор как этот столь отдаленный период Земля, несомненно, претерпела множество великих революций, и было достаточно времени для миграции в самых больших масштабах.
— Чарльз Дарвин, Происхождение человека[86]
В 1871 году человеческие окаменелости древних гомининов почти не были доступны. Почти пятьдесят лет спустя предположение Дарвина подтвердилось, когда антропологи начали находить окаменелости древних гомининов с маленьким мозгом в нескольких районах Африки (список окаменелостей гоминин ). Гипотеза недавний (в отличие от архаичный ) Африканское происхождение развилось в 20 веке. «Недавнее африканское происхождение» современных людей означает «единственное происхождение» (моногенизм) и использовалось в различных контекстах как антоним полигенизма. К середине ХХ века дебаты в антропологии склонились в пользу моногенизма. Отдельные сторонники полигенизма середины 20 века, такие как Карлтон Кун, который еще в 1962 году думал, что Х. сапиенс возник пять раз из H. erectus в пяти местах.[87]
Гипотеза о межрегиональном происхождении
Исторической альтернативой недавней модели происхождения является мультирегиональное происхождение современного человека, первоначально предложенный Милфорд Уолпофф в 1980-е гг. Эта точка зрения предполагает, что происхождение анатомически современных человеческих популяций от H. erectus в начале Плейстоцен 1,8 миллиона лет назад произошло в пределах постоянного населения мира. Гипотеза обязательно отвергает предположение барьер бесплодия между древними евразийскими и африканскими популяциями Гомо. В конце 1980-х и 1990-х годах эта гипотеза вызывала споры.[88] Нынешняя терминология «недавнего происхождения» и «из Африки» стала актуальной в контексте этих дебатов в 1990-х годах.[89] Первоначально рассматриваемая как противоположная альтернатива модели недавнего происхождения, мультирегиональная гипотеза в ее первоначальной «сильной» форме устарела, в то время как ее различные модифицированные более слабые варианты стали вариантами взгляда на «недавнее происхождение» в сочетании с архаичная примесь.[90] Стрингер (2014) различает исходную или «классическую» межрегиональную модель, существовавшую с 1984 года (ее формулировка) до 2003 года, от «слабого» варианта после 2003 года, который «приблизился к модели ассимиляции».[91][92]
Митохондриальные анализы
В 1980-х годах Аллан Уилсон вместе с Ребекка Л. Канн и Марк Стоункинг работал над генетическим датированием последнего по материнской линии общего предка современных человеческих популяций (получившего название "Митохондриальная Ева "). Для обозначения информативных генетические маркеры для отслеживания истории эволюции человека Уилсон сосредоточился на митохондриальная ДНК (мтДНК) передается от матери к ребенку. Этот материал ДНК быстро мутирует, что позволяет легко отображать изменения за относительно короткое время. Своим открытием, что человеческая мтДНК генетически гораздо менее разнообразна, чем мтДНК шимпанзе, Уилсон пришел к выводу, что современные человеческие популяции недавно расходились от одной популяции, в то время как более старые человеческие виды, такие как Неандертальцы и человек прямоходящий вымерли.[93] С появлением археогенетика в 1990-е годы датирование митохондрий и Y-хромосомные гаплогруппы стало возможным с некоторой уверенностью. К 1999 г. оценка составляла около 150 000 лет для mt-MRCA и от 60 000 до 70 000 лет для миграции из Африки.[94]
С 2000 по 2003 год велись споры о митохондриальной ДНК "Манго Человек 3 "(LM3) и его возможное влияние на мультирегиональную гипотезу. Было обнаружено, что LM3 имеет большее, чем ожидалось, количество различий в последовательностях по сравнению с современной ДНК человека (CRS ).[95] Сравнение митохондриальной ДНК с ДНК древних и современных аборигенов привело к выводу, что человек манго выходит за пределы диапазона генетических вариаций, наблюдаемых у австралийских аборигенов, и был использован для подтверждения гипотезы о мультирегиональном происхождении. Повторный анализ LM3 и других древних образцов из этого района, опубликованный в 2016 году, показал, что он схож с последовательностями современных аборигенов Австралии, что противоречит результатам более раннего исследования.[96]
Y-хромосома анализы
Согласно текущим оценкам Самый последний общий предок мужского пола ("Y-хромосомный Адам "или Y-MRCA) сходятся с оценками возраста анатомически современные люди и задолго до миграции из Африки, гипотезы о географическом происхождении продолжают ограничиваться Африканский континент.
Наиболее базальные линии были обнаружены в Запад, Северо-Запад и Центральная Африка, предполагая правдоподобие Y-MRCA, проживающего в основном регионе «Центрально-Северо-Западная Африка».[97]
другое исследование находит правдоподобное положение в «северо-западном квадранте африканского континента» для возникновения гаплогруппы A1b.[98] Отчет 2013 г. гаплогруппа A00 найден среди Мбо люди современного западного Камерун также совместим с этим изображением.[99]
Пересмотр филогении Y-хромосомы с 2011 г. повлиял на оценки вероятного географического происхождения Y-MRCA, а также на оценки временной глубины. По той же причине будущее открытие неизвестных в настоящее время архаических гаплогрупп у живых людей снова приведет к таким пересмотрам. В частности, возможное присутствие от 1% до 4% ДНК неандертальца в евразийских геномах подразумевает, что (маловероятное) событие обнаружения единственного живого евразийского самца, демонстрирующего неандертальскую патрилинейную линию, немедленно отодвинет T-MRCA («время до MRCA») как минимум вдвое по сравнению с текущей оценкой. Однако открытие Y-хромосомы неандертальца Мендесом и другие. Исследование, проведенное в 2016 году, предполагает исчезновение неандертальских отцов, поскольку происхождение, выведенное из последовательности неандертальцев, выходит за рамки диапазона современный человек генетическая изменчивость.[100] Вопросы географического происхождения станут частью дискуссии об эволюции неандертальцев с человек прямоходящий.
Смотрите также
- Археогенетика Ближнего Востока
- Поведенческая современность - Переход человеческого вида к антропологически современному поведению
- Прибрежная миграция
- Рассвет человечества (Фильм PBS 2015 года)
- Ранние миграции людей - Распространение людей из Африки по миру
- Генетика и археогенетика Южной Азии
- Генетическая история Европы
- Генетическая история коренных народов Америки
- Генетическая история Италии
- Генетическая история Северной Африки
- Генетическая история Британских островов
- Генетическая история Пиренейского полуострова
- Череп Хофмейра
- Эволюция человека - Эволюционный процесс, ведущий к анатомически современным людям
- Человеческое происхождение
- Хронология человека - События гомининов за последние 10 миллионов лет
- Точка идентичных предков
- Теория индоарийской миграции
- Теория насоса Сахары
- Невероятное человеческое путешествие
- Хронология эволюции человека - Хронологический очерк основных событий в развитии человеческого вида
Примечания
- ^ С 1984 по 2003 год альтернативной научной гипотезой была мультирегиональное происхождение современного человека, который предвидел волну Homo sapiens ранее мигрировавшие из Африки и скрещивание с местными человек прямоходящий населения в различных регионах земного шара.Джермейн Р., Килгор Л., Треватан В. (2008). Основы физической антропологии. Cengage Learning. С. 266–. ISBN 978-0-495-50939-4. Получено 14 июн 2011.
- ^ Макчесни (2015): "... генетические данные свидетельствуют о том, что небольшая группа с маркером M168 мигрировала из Африки вдоль побережья Аравийского полуострова и Индии через Индонезию и достигла Австралии очень рано, между 60 000 и 50 000 лет назад. Это очень ранняя миграция. в Австралию также поддерживается Расмуссеном и др. (2011) ".
- ^ Макчесни (2015): «Уэллс (2003) разделил потомков мужчин, покинувших Африку, на генеалогическое древо с 11 линиями. Каждый генетический маркер представляет собой одноточечную мутацию (SNP) в определенном месте генома. Во-первых, генетические данные свидетельствуют о том, что небольшая полоса с маркером M168 мигрировала из Африки вдоль побережья Аравийского полуострова и Индии через Индонезию и достигла Австралии очень рано, между 60 000 и 50 000 лет назад. Эта очень ранняя миграция в Австралию также подтверждается Расмуссеном и др. ( 2011). Во-вторых, группа с маркером M89 вышла из северо-востока Африки на Ближний Восток 45 000 лет назад. Оттуда группа M89 разделилась на две группы. Одна группа, разработавшая маркер M9, ушла в Азию около 40 000 лет назад. Азиатская группа (M9) разделилась тремя путями: на Центральную Азию (M45), 35 000 лет назад; на Индию (M20), 30 000 лет назад; и на Китай (M122), 10 000 лет назад. Центральноазиатская группа (M45) разделилась на две группы: в сторону Европы (M173), 3 0 000 лет назад и в сторону Сибири (M242), 20 000 лет назад. Наконец, сибирская группа (M242) продолжила заселять Северную и Южную Америку (M3) около 10 000 лет назад.[26]
- ^ Исследователи использовали методы радиоуглеродного датирования пыльцевых зерен, пойманных в иле на дне озера, чтобы установить растительность на протяжении многих лет. Малави озеро в Африке, образцы берутся каждые 300 лет. В образцах, взятых во время сильной засухи, было мало пыльцы или древесного угля, что свидетельствует о скудной растительности, которую можно было почти не сжигать. Территория вокруг Озеро Малави сегодня густо засаженный деревьями, приблизительно 135 000–90 000 лет назад был пустыней.[39]
Рекомендации
- ^ Лю, Пругнолле и др. (2006). «Доступные в настоящее время генетические и археологические данные подтверждают недавнее единственное происхождение современных людей в Восточной Африке. Однако на этом консенсус по истории человеческих поселений заканчивается, и значительная неопределенность омрачает любые более подробные аспекты истории человеческой колонизации».
- ^ Стрингер С. (июнь 2003 г.). «Эволюция человека: из Эфиопии». Природа. 423 (6941): 692–3, 695. Bibcode:2003Натура.423..692S. Дои:10.1038 / 423692a. PMID 12802315.
- ^ Стрингер C (13 марта 2012 г.). Одинокие выжившие: как мы стали единственными людьми на Земле. Генри Холт и компания. п. 26. ISBN 978-1-4299-7344-1.
- ^ Wolpoff MH, Hawks J, Caspari R (май 2000 г.). «Мультирегиональное, а не множественное происхождение» (PDF). Американский журнал физической антропологии. 112 (1): 129–36. Дои:10.1002 / (SICI) 1096-8644 (200005) 112: 1 <129 :: AID-AJPA11> 3.0.CO; 2-K. HDL:2027.42/34270. PMID 10766948.
- ^ Mafessoni F (январь 2019). «Встречи с архаичными гомининами». Природа Экология и эволюция. 3 (1): 14–15. Дои:10.1038 / s41559-018-0729-6. PMID 30478304.
- ^ Villanea FA, Schraiber JG (январь 2019 г.). «Множественные эпизоды скрещивания неандертальцев и современных людей». Природа Экология и эволюция. 3 (1): 39–44. Дои:10.1038 / с41559-018-0735-8. ЧВК 6309227. PMID 30478305.
- ^ а б Университет Хаддерсфилда (20 марта 2019 г.). «Исследователи пролили новый свет на происхождение современного человека - работа, опубликованная в журнале Nature, подтверждает, что расселение Homo sapiens с юга на восток Африки непосредственно предшествовало миграции из Африки». EurekAlert!. Получено 23 марта 2019.
- ^ а б Рито Т., Виейра Д., Сильва М., Конде-Соуза Е., Перейра Л., Мелларс П. и др. (Март 2019 г.). «Расселение Homo sapiens с юга на восток Африки непосредственно предшествовало миграции из Африки». Научные отчеты. 9 (1): 4728. Bibcode:2019НатСР ... 9.4728R. Дои:10.1038 / с41598-019-41176-3. ЧВК 6426877. PMID 30894612.
- ^ Циммер С (10 июля 2019 г.). «Кость черепа, обнаруженная в Греции, может изменить предысторию человеческой предыстории - кость, найденная в пещере, является старейшей современной окаменелостью человека, когда-либо обнаруженной в Европе. Это намекает на то, что люди начали покидать Африку намного раньше, чем считалось ранее».. Нью-Йорк Таймс. Получено 11 июля 2019.
- ^ Персонал (10 июля 2019 г.). "'Самые старые останки 'за пределами Африки сбрасывают часы миграции людей ". Phys.org. Получено 10 июля 2019.
- ^ Харвати К., Рёдинг С., Босман А.М., Каракостис Ф.А., Грюн Р., Стрингер С. и др. (Июль 2019). «Окаменелости пещеры Апидима являются самым ранним свидетельством существования Homo sapiens в Евразии». Природа. 571 (7766): 500–504. Дои:10.1038 / с41586-019-1376-з. PMID 31292546.
- ^ а б c Armitage SJ, Jasim SA, Marks AE, Parker AG, Usik VI, Uerpmann HP (январь 2011 г.). «Южный путь« из Африки »: свидетельство ранней экспансии современного человека в Аравию». Наука. 331 (6016): 453–6. Bibcode:2011Наука ... 331..453А. Дои:10.1126 / science.1199113. PMID 21273486.
- ^ а б c Балтер М (январь 2011 г.). «Была ли Северная Африка стартовой площадкой для современных миграций людей?» (PDF). Наука. 331 (6013): 20–3. Bibcode:2011Научный ... 331 ... 20B. Дои:10.1126 / science.331.6013.20. PMID 21212332.
- ^ а б Cruciani F, Trombetta B, Massaia A, Destro-Bisol G, Sellitto D, Scozzari R (июнь 2011 г.). "Обновленный корень филогенетического дерева Y-хромосомы человека: происхождение отцовского разнообразия в Африке". Американский журнал генетики человека. 88 (6): 814–818. Дои:10.1016 / j.ajhg.2011.05.002. ЧВК 3113241. PMID 21601174.
- ^ а б Smith TM, Tafforeau P, Reid DJ, Grün R, Eggins S, Boutakiout M, Hublin JJ (апрель 2007 г.). «Самые ранние свидетельства современной истории человеческой жизни в Северной Африке ранних Homo sapiens». Труды Национальной академии наук Соединенных Штатов Америки. 104 (15): 6128–33. Bibcode:2007ПНАС..104.6128С. Дои:10.1073 / pnas.0700747104. ЧВК 1828706. PMID 17372199.
- ^ Bae CJ, Douka K, Petraglia MD (декабрь 2017 г.). «О происхождении современного человека: азиатские перспективы». Наука. 358 (6368): eaai9067. Дои:10.1126/science.aai9067. PMID 29217544.
- ^ Kuo L (10 December 2017). "Early humans migrated out of Africa much earlier than we thought". Кварцевый. Получено 10 декабря 2017.
- ^ а б Liu, Martinón-Torres et al. (2015).
Смотрите также Modern humans in China ~80,000 years ago (?), Dieneks' Anthropology Blog. - ^ Posth C, Renaud G, Mittnik M, Drucker DG, Rougier H, Cupillard C, et al. (2016). "Pleistocene Mitochondrial Genomes Suggest a Single Major Dispersal of Non-Africans and a Late Glacial Population Turnover in Europe". Текущая биология. 26 (6): 827–833. Дои:10.1016/j.cub.2016.01.037. HDL:2440/114930. PMID 26853362.
- ^ Кармин М., Сааг Л., Висенте М., Вильсон Сайрес М.А., Ярве М., Талас Ю.Г. и др. (April 2015). «Недавнее узкое место разнообразия Y-хромосомы совпадает с глобальным изменением культуры». Геномные исследования. 25 (4): 459–66. Дои:10.1101 / гр.186684.114. ЧВК 4381518. PMID 25770088.
- ^ а б c Haber M, Jones AL, Connell BA, Arciero E, Yang H, Thomas MG, et al. (August 2019). "A Rare Deep-Rooting D0 African Y-Chromosomal Haplogroup and Its Implications for the Expansion of Modern Humans Out of Africa". Генетика. 212 (4): 1421–1428. Дои:10.1534/genetics.119.302368. ЧВК 6707464. PMID 31196864.
- ^ а б Clarkson C, Jacobs Z, Marwick B, Fullagar R, Wallis L, Smith M, et al. (Июль 2017 г.). "Human occupation of northern Australia by 65,000 years ago". Природа. 547 (7663): 306–310. Bibcode:2017Натура.547..306C. Дои:10.1038 / природа22968. HDL:2440/107043. PMID 28726833.
- ^ а б c St Fleu N (19 July 2017). "Humans First Arrived in Australia 65,000 Years Ago, Study Suggests". Нью-Йорк Таймс.
- ^ а б c Wood R (2 September 2017). "Comments on the chronology of Madjedbebe". Австралийская археология. 83 (3): 172–174. Дои:10.1080/03122417.2017.1408545. ISSN 0312-2417.
- ^ а б c O'Connell JF, Allen J, Williams MA, Williams AN, Turney CS, Spooner NA, et al. (Август 2018 г.). "Homo sapiens first reach Southeast Asia and Sahul?". Труды Национальной академии наук Соединенных Штатов Америки. 115 (34): 8482–8490. Дои:10.1073/pnas.1808385115. ЧВК 6112744. PMID 30082377.
- ^ а б c McChesney 2015.
- ^ а б c d е Macaulay et al. (2005).
- ^ а б Posth et al. (2016).
Смотрите также mtDNA from 55 hunter-gatherers across 35,000 years in Europe, Dienekes' Anthroplogy Bog. - ^ Prüfer K, Racimo F, Patterson N, Jay F, Sankararaman S, Sawyer S, et al. (January 2014) [Online 2013]. «Полная последовательность генома неандертальца с Горного Алтая». Природа. 505 (7481): 43–9. Bibcode:2014Натура.505 ... 43П. Дои:10.1038 / природа12886. ЧВК 4031459. PMID 24352235.
- ^ а б Lachance J, Vernot B, Elbers CC, Ferwerda B, Froment A, Bodo JM, et al. (Август 2012 г.). "Evolutionary history and adaptation from high-coverage whole-genome sequences of diverse African hunter-gatherers". Клетка. 150 (3): 457–69. Дои:10.1016/j.cell.2012.07.009. ЧВК 3426505. PMID 22840920.
- ^ а б Hammer MF, Woerner AE, Mendez FL, Watkins JC, Wall JD (September 2011). "Genetic evidence for archaic admixture in Africa". Труды Национальной академии наук Соединенных Штатов Америки. 108 (37): 15123–8. Bibcode:2011PNAS..10815123H. Дои:10.1073/pnas.1109300108. ЧВК 3174671. PMID 21896735.
- ^ а б c Douka K, Bergman CA, Hedges RE, Wesselingh FP, Higham TF (11 September 2013). "Chronology of Ksar Akil (Lebanon) and implications for the colonization of Europe by anatomically modern humans". PLOS One. 8 (9): e72931. Bibcode:2013PLoSO...872931D. Дои:10.1371/journal.pone.0072931. ЧВК 3770606. PMID 24039825.
- ^ Hammond, Ashley S.; Royer, Danielle F.; Fleagle, John G. (July 2017). "The Omo-Kibish I pelvis". Журнал эволюции человека. 108: 199–219. Дои:10.1016/j.jhevol.2017.04.004. ISSN 1095-8606. PMID 28552208.
- ^ а б c d Beyin (2011).
- ^ Posth C, Wißing C, Kitagawa K, Pagani L, van Holstein L, Racimo F, et al. (Июль 2017 г.). "Deeply divergent archaic mitochondrial genome provides lower time boundary for African gene flow into Neanderthals". Nature Communications. 8: 16046. Bibcode:2017NatCo...816046P. Дои:10.1038/ncomms16046. ЧВК 5500885. PMID 28675384.; смотрите также Zimmer C (4 July 2017). "In Neanderthal DNA, Signs of a Mysterious Human Migration". Нью-Йорк Таймс. Получено 4 июля 2017..
- ^ "Scientists discover oldest known modern human fossil outside of Africa: Analysis of fossil suggests Homo sapiens left Africa at least 50,000 years earlier than previously thought". ScienceDaily. Получено 27 января 2018.
- ^ Ghosh P (2018). "Modern humans left Africa much earlier". Новости BBC. Получено 27 января 2018.
- ^ а б c d е Appenzeller (2012).
- ^ а б Jensen MN (8 October 2007). "Newfound Ancient African Megadroughts May Have Driven Evolution of Humans and Fish. The findings provide new insights into humans' migration out of Africa and the evolution of fishes in Africa's Great Lakes". Университет Аризоны. Получено 25 сентября 2017.
- ^ Finlayson (2009), п. 68.
- ^ Liu, Prugnolle et al. (2006).
- ^ Scerri EM, Drake NA, Jennings R, Groucutt HS (1 October 2014). "Earliest evidence for the structure of Homo sapiens populations in Africa". Четвертичные научные обзоры. 101: 207–216. Bibcode:2014QSRv..101..207S. Дои:10.1016/j.quascirev.2014.07.019.
- ^ Rose JI, Usik VI, Marks AE, Hilbert YH, Galletti CS, Parton A, et al. (2011). "The Nubian Complex of Dhofar, Oman: an African middle Stone Age industry in Southern Arabia". PLOS One. 6 (11): e28239. Bibcode:2011PLoSO...628239R. Дои:10.1371/journal.pone.0028239. ЧВК 3227647. PMID 22140561.
- ^ а б c Kuhlwilm et al. (2016).
Смотрите также Ancestors of Eastern Neandertals admixed with modern humans 100 thousand years ago, Dienekes'Anthropology Blog. - ^ Gibbons, Ann (2 March 2017). "Ancient skulls may belong to elusive humans called Denisovans". Наука. Дои:10.1126/science.aal0846. Получено 25 сентября 2017.
- ^ Shen G, Wang W, Wang Q, Zhao J, Collerson K, Zhou C, Tobias PV (December 2002). «U-серия датировки сайта гоминидов Люцзян в Гуанси, Южный Китай». Журнал эволюции человека. 43 (6): 817–29. Дои:10.1006 / jhev.2002.0601. PMID 12473485.
- ^ а б Rosenburg K (2002). "A Late Pleistocene Human Skeleton from Liujiang, China Suggests Regional Population Variation in Sexual Dimorphism in the Human Pelvis". Variability and Evolution.
- ^ Kaifu Y (2012). "Fossil Record of Early Modern Humans in East Asia". Palaeoenvironmental Changes and Human Dispersals in North and East Asia During MIS3 and MIS2. 248: 2–11. Bibcode:2012QuInt.248....2K. Дои:10.1016/j.quaint.2011.02.017.
- ^ Zhivotovsky LA, Rosenberg NA, Feldman MW (May 2003). "Features of evolution and expansion of modern humans, inferred from genomewide microsatellite markers". Американский журнал генетики человека. 72 (5): 1171–86. Дои:10.1086/375120. ЧВК 1180270. PMID 12690579.
- ^ Stix G (2008). "The Migration History of Humans: DNA Study Traces Human Origins Across the Continents". Получено 14 июн 2011.
- ^ а б c Metspalu M, Kivisild T, Metspalu E, Parik J, Hudjashov G, Kaldma K, et al. (Август 2004 г.). "Most of the extant mtDNA boundaries in south and southwest Asia were likely shaped during the initial settlement of Eurasia by anatomically modern humans". BMC Genetics. 5: 26. Дои:10.1186/1471-2156-5-26. ЧВК 516768. PMID 15339343.
- ^ Fernandes CA, Rohling EJ, Siddall M (June 2006). "Absence of post-Miocene Red Sea land bridges: biogeographic implications". Журнал биогеографии. 33 (6): 961–66. Дои:10.1111/j.1365-2699.2006.01478.x.
- ^ Walter RC, Buffler RT, Bruggemann JH, Guillaume MM, Berhe SM, Negassi B, et al. (Май 2000 г.). "Early human occupation of the Red Sea coast of Eritrea during the last interglacial". Природа. 405 (6782): 65–9. Bibcode:2000Natur.405...65W. Дои:10.1038/35011048. PMID 10811218.
- ^ Catherine B (24 November 2012). "Our True Dawn". Новый ученый (2892): 34–37. ISSN 0262-4079.
- ^ Karmin M, Saag L, Vicente M, Wilson Sayres MA, Järve M, et al. (April 2015). «Недавнее узкое место разнообразия Y-хромосомы совпадает с глобальным изменением культуры». Геномные исследования. 25 (4): 459–66. Дои:10.1101 / гр.186684.114. ЧВК 4381518. PMID 25770088.
- ^ а б Vai S, Sarno S, Lari M, Luiselli D, Manzi G, Gallinaro M, et al. (Март 2019 г.). "Ancestral mitochondrial N lineage from the Neolithic 'green' Sahara". Научные отчеты. 9 (1): 3530. Bibcode:2019NatSR...9.3530V. Дои:10.1038/s41598-019-39802-1. ЧВК 6401177. PMID 30837540.
- ^ Hershkovitz et al. (2015)
Смотрите также 55,000-Year-Old Skull Fossil Sheds New Light on Human Migration out of Africa, Science News. - ^ Bowler JM, Johnston H, Olley JM, Prescott JR, Roberts RG, Shawcross W, Spooner NA (February 2003). "New ages for human occupation and climatic change at Lake Mungo, Australia". Природа. 421 (6925): 837–40. Bibcode:2003Natur.421..837B. Дои:10.1038/nature01383. PMID 12594511.
- ^ Оллея Дж. М., Робертс Р. Г., Йошида Х., Боулер Дж. М. (2006). «Однозерновое оптическое датирование могилы-засыпки, связанной с человеческими захоронениями на озере Мунго, Австралия». Четвертичные научные обзоры. 25 (19–20): 2469–74. Bibcode:2006QSRv ... 25.2469O. Дои:10.1016 / j.quascirev.2005.07.022.
- ^ "A relative from the Tianyuan Cave". Общество Макса Планка. 21 января 2013 г.
- ^ "A relative from the Tianyuan Cave: Humans living 40,000 years ago likely related to many present-day Asians and Native Americans". Science Daily. 21 января 2013 г.
- ^ "DNA Analysis Reveals Common Origin of Tianyuan Humans and Native Americans, Asians". Научные новости. 24 января 2013 г.
- ^ "Ancient human DNA suggests minimal interbreeding". Новости науки. 21 января 2013 г.
- ^ "Ancient Bone DNA Shows Ancestry of Modern Asians & Native Americans". Новости пещер. 31 января 2013 г.
- ^ Hu Y, Shang H, Tong H, Nehlich O, Liu W, Zhao C, et al. (Июль 2009 г.). "Stable isotope dietary analysis of the Tianyuan 1 early modern human". Труды Национальной академии наук Соединенных Штатов Америки. 106 (27): 10971–4. Bibcode:2009PNAS..10610971H. Дои:10.1073/pnas.0904826106. ЧВК 2706269. PMID 19581579.
- ^ Brown P (August 1992). «Недавняя эволюция человека в Восточной Азии и Австралазии». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 337 (1280): 235–42. Bibcode:1992RSPTB.337..235B. Дои:10.1098 / rstb.1992.0101. PMID 1357698.
- ^ Gonder MK, Mortensen HM, Reed FA, de Sousa A, Tishkoff SA (March 2007). "Whole-mtDNA genome sequence analysis of ancient African lineages". Молекулярная биология и эволюция. 24 (3): 757–68. Дои:10.1093/molbev/msl209. PMID 17194802.
- ^ Chen YS, Olckers A, Schurr TG, Kogelnik AM, Huoponen K, Wallace DC (April 2000). "mtDNA variation in the South African Kung and Khwe-and their genetic relationships to other African populations". Американский журнал генетики человека. 66 (4): 1362–83. Дои:10.1086/302848. ЧВК 1288201. PMID 10739760.
- ^ Soares P, Alshamali F, Pereira JB, Fernandes V, Silva NM, Afonso C, et al. (Март 2012 г.). "The Expansion of mtDNA Haplogroup L3 within and out of Africa". Молекулярная биология и эволюция. 29 (3): 915–27. Дои:10.1093/molbev/msr245. PMID 22096215.
- ^ Groucutt et al. (2015).
- ^ Maca-Meyer N, González AM, Larruga JM, Flores C, Cabrera VM (13 August 2001). «Основные геномные митохондриальные линии определяют раннюю экспансию человека». BMC Genetics. 2: 13. Дои:10.1186/1471-2156-2-13. ЧВК 55343. PMID 11553319.
- ^ Ingman M, Gyllensten U (июль 2003 г.). «Вариации митохондриального генома и история эволюции аборигенов Австралии и Новой Гвинеи». Геномные исследования. 13 (7): 1600–6. Дои:10.1101 / гр.686603. ЧВК 403733. PMID 12840039.
- ^ Yu N, Chen FC, Ota S, Jorde LB, Pamilo P, Patthy L, et al. (Май 2002 г.). "Larger genetic differences within africans than between Africans and Eurasians". Генетика. 161 (1): 269–74. ЧВК 1462113. PMID 12019240. Получено 7 апреля 2013.
- ^ Coop G, Pickrell JK, Novembre J, Kudaravalli S, Li J, Absher D, et al. (Июнь 2009 г.). Schierup MH (ed.). "The role of geography in human adaptation". PLOS Genetics. 5 (6): e1000500. Дои:10.1371/journal.pgen.1000500. ЧВК 2685456. PMID 19503611.;summary in Kliman (ed.), Encyclopedia of Evolutionary Biology (2016), п. 451
- ^ Gurdasani D, Carstensen T, Tekola-Ayele F, Pagani L, Tachmazidou I, Hatzikotoulas K, et al. (Январь 2015 г.). "The African Genome Variation Project shapes medical genetics in Africa". Природа. 517 (7534): 327–32. Bibcode:2015Natur.517..327G. Дои:10.1038/nature13997. ЧВК 4297536. PMID 25470054.
- ^ Matisoo-Smith E, Horsburgh KA (2016). ДНК для археологов. Рутледж.
- ^ Rasmussen M, Guo X, Wang Y, Lohmueller KE, Rasmussen S, Albrechtsen A, et al. (Октябрь 2011 г.). "An Aboriginal Australian genome reveals separate human dispersals into Asia". Наука. 334 (6052): 94–8. Bibcode:2011Sci...334...94R. Дои:10.1126/science.1211177. ЧВК 3991479. PMID 21940856.
- ^ East Asians 2.3–2.6%, Western Eurasians 1.8–2.4% (Prüfer K, de Filippo C, Grote S, Mafessoni F, Korlević P, Hajdinjak M, et al. (Ноябрь 2017 г.). "A high-coverage Neandertal genome from Vindija Cave in Croatia". Наука. 358 (6363): 655–658. Bibcode:2017Sci...358..655P. Дои:10.1126/science.aao1887. ЧВК 6185897. PMID 28982794.)
- ^ Callaway E (2012). "Hunter-gatherer genomes a trove of genetic diversity". Природа. Дои:10.1038/nature.2012.11076.
- ^ Xu D, Pavlidis P, Taskent RO, Alachiotis N, Flanagan C, DeGiorgio M, et al. (Октябрь 2017 г.). "Archaic Hominin Introgression in Africa Contributes to Functional Salivary MUC7 Genetic Variation". Молекулярная биология и эволюция. 34 (10): 2704–2715. Дои:10.1093/molbev/msx206. ЧВК 5850612. PMID 28957509.
- ^ Petraglia M, Clarkson C, Boivin N, Haslam M, Korisettar R, Chaubey G, et al. (Июль 2009 г.). "Population increase and environmental deterioration correspond with microlithic innovations in South Asia ca. 35,000 years ago". Труды Национальной академии наук Соединенных Штатов Америки. 106 (30): 12261–6. Bibcode:2009ПНАС..10612261П. Дои:10.1073 / pnas.0810842106. ЧВК 2718386. PMID 19620737.
- ^ Lafreniere P (2010). Adaptive Origins: Evolution and Human Development. Тейлор и Фрэнсис. п. 90. ISBN 978-0-8058-6012-2. Получено 14 июн 2011.
- ^ Robinson D, Ash PM (2010). The Emergence of Humans: An Exploration of the Evolutionary Timeline. Нью-Йорк: Вили. ISBN 978-0-470-01315-1.
- ^ Palmer D (2006). Prehistoric Past Revealed: The Four Billion Year History of Life on Earth. Беркли: Калифорнийский университет Press. п. 43. ISBN 978-0-520-24827-4.
- ^ Regal B (2004). Human evolution: a guide to the debates. Санта-Барбара, Калифорния: ABC-CLIO. С. 73–75. ISBN 978-1-85109-418-9.
- ^ "The descent of man Chapter 6 – On the Affinities and Genealogy of Man". Darwin-online.org.uk. Получено 11 января 2011.
- ^ Jackson JP Jr (2001). "'In Ways Unacademical': The Reception of Carleton S. Coon's The Origin of Races" (PDF). Журнал истории биологии. 34 (2): 247–85. Дои:10.1023/A:1010366015968. Архивировано из оригинал (PDF) 14 мая 2013 г.
- ^ Stringer CB, Andrews P (March 1988). "Genetic and fossil evidence for the origin of modern humans". Наука. 239 (4845): 1263–8. Bibcode:1988Sci...239.1263S. Дои:10.1126/science.3125610. PMID 3125610.Stringer C, Bräuer G (1994). "Methods, misreading, and bias". Американский антрополог. 96 (2): 416–24. Дои:10.1525/aa.1994.96.2.02a00080.
Stringer CB (1992). "Replacement, continuity and the origin of Homo sapiens". In Smith FH (ed.). Continuity or replacement? Controversies in Homo sapiens evolution. Роттердам: Балкема. pp. 9–24.
Bräuer G, Stringer C (1997). "Models, polarization, and perspectives on modern human origins". Conceptual issues in modern human origins research. Нью-Йорк: Альдин де Грюйтер. pp. 191–201. - ^ Wu L (1997). "The dental continuity of humans in China from Pleistocene to Holocene, and the origin of mongoloids". Quaternary Geology. 21: 24–32. ISBN 978-90-6764-243-9.
- ^ Stringer C (2001). "Modern human origins – distinguishing the models". Afr. Археол. Rev. 18 (2): 67–75. Дои:10.1023/A:1011079908461.
- ^ Stringer C (April 2002). "Modern human origins: progress and prospects". Философские труды Лондонского королевского общества. Серия B, Биологические науки. 357 (1420): 563–79. Дои:10.1098/rstb.2001.1057. ЧВК 1692961. PMID 12028792.
- ^ Stringer C (May 2014). "Why we are not all multiregionalists now". Тенденции в экологии и эволюции. 29 (5): 248–51. Дои:10.1016/j.tree.2014.03.001. PMID 24702983.
- ^ "Allan Wilson: Revolutionary Evolutionist". New Zealanders Heroes.
- ^ Wallace DC, Brown MD, Lott MT (September 1999). "Mitochondrial DNA variation in human evolution and disease". Ген. 238 (1): 211–30. Дои:10.1016/S0378-1119(99)00295-4. PMID 10570998. "evidence that our species arose in Africa about 150 000 years before present (YBP), migrated out of Africa into Asia about 60 000 to 70 000 YBP and into Europe about 40 000 to 50 000 YBP, and migrated from Asia and possibly Europe to the Americas about 20 000 to 30 000 YBP."
- ^ Adcock GJ, Dennis ES, Easteal S, Huttley GA, Jermiin LS, Peacock WJ, Thorne A (January 2001). «Последовательности митохондриальной ДНК у древних австралийцев: значение для современного происхождения человека». Труды Национальной академии наук Соединенных Штатов Америки. 98 (2): 537–42. Bibcode:2001ПНАС ... 98..537А. Дои:10.1073 / пнас.98.2.537. ЧВК 14622. PMID 11209053.
- ^ Heupink TH, Subramanian S, Wright JL, Endicott P, Westaway MC, Huynen L, et al. (Июнь 2016). «Возвращение к древним последовательностям мтДНК первых австралийцев». Труды Национальной академии наук Соединенных Штатов Америки. 113 (25): 6892–7. Дои:10.1073 / pnas.1521066113. ЧВК 4922152. PMID 27274055.
- ^ Cruciani, Fulvio; Trombetta, Beniamino; Massaia, Andrea; Дестро-Бизоль, Джованни; Sellitto, Daniele; Scozzari, Rosaria (2011). "A Revised Root for the Human Y Chromosomal Phylogenetic Tree: The Origin of Patrilineal Diversity in Africa". Американский журнал генетики человека. 88 (6): 814–18. Дои:10.1016 / j.ajhg.2011.05.002. ЧВК 3113241. PMID 21601174.
- ^ Scozzari R; Massaia A; D'Atanasio E; Myres NM; Perego UA; и другие. (2012). Карамелли, Дэвид (ред.). "Molecular Dissection of the Basal Clades in the Human Y Chromosome Phylogenetic Tree". PLOS One. 7 (11): e49170. Bibcode:2012PLoSO ... 749170S. Дои:10.1371 / journal.pone.0049170. ЧВК 3492319. PMID 23145109.
- ^ Mendez, Fernando; Krahn, Thomas; Schrack, Bonnie; Krahn, Astrid-Maria; Veeramah, Krishna; Woerner, August; Fomine, Forka Leypey Mathew; Брэдман, Нил; Thomas, Mark; Карафет, Татьяна М .; Hammer, Michael F. (7 March 2013). "An African American paternal lineage adds an extremely ancient root to the human Y chromosome phylogenetic tree" (PDF). Американский журнал генетики человека. 92 (3): 454–59. Дои:10.1016/j.ajhg.2013.02.002. ЧВК 3591855. PMID 23453668.
- ^ Mendez, L.; и другие. (2016). "Дивергенция Y-хромосом неандертальца и современного человека". Американский журнал генетики человека. 98 (4): 728–34. Дои:10.1016 / j.ajhg.2016.02.023. ЧВК 4833433. PMID 27058445.
Источники
- Appenzeller T (2012). "Human migrations: Eastern odyssey. Humans had spread across Asia by 50,000 years ago. Everything else about our original exodus from Africa is up for debate". Природа. 485 (7396).
- Beyin A (2011). «Расселение людей из Африки в верхнем плейстоцене: обзор текущего состояния дебатов». International Journal of Evolutionary Biology. 2011 (615094): 615094. Дои:10.4061/2011/615094. ЧВК 3119552. PMID 21716744.
- Endicott P, Gilbert MT, Stringer C, Lalueza-Fox C, Willerslev E, Hansen AJ, Cooper A (January 2003). "The genetic origins of the Andaman Islanders". Американский журнал генетики человека. 72 (1): 178–84. Дои:10.1086/345487. ЧВК 378623. PMID 12478481.
- Finlayson C (2009). The humans who went extinct: why Neanderthals died out and we survived. Oxford University Press, США. ISBN 978-0-19-923918-4.
- Groucutt HS, Petraglia MD, Bailey G, Scerri EM, Parton A, Clark-Balzan L, et al. (2015). "Rethinking the dispersal of Homo sapiens out of Africa". Эволюционная антропология. 24 (4): 149–64. Дои:10.1002/evan.21455. ЧВК 6715448. PMID 26267436.
- Harding RM, Healy E, Ray AJ, Ellis NS, Flanagan N, Todd C, et al. (Апрель 2000 г.). "Evidence for variable selective pressures at MC1R". Американский журнал генетики человека. 66 (4): 1351–61. Дои:10.1086/302863. ЧВК 1288200. PMID 10733465.
- Hershkovitz I, Marder O, Ayalon A, Bar-Matthews M, Yasur G, Boaretto E, et al. (April 2015). «Левантийский череп из пещеры Манот (Израиль) предвещает появление первых современных людей в Европе». Природа. 520 (7546): 216–9. Bibcode:2015Natur.520..216H. Дои:10.1038 / природа14134. PMID 25629628.
- Kuhlwilm M, Gronau I, Hubisz MJ, de Filippo C, Prado-Martinez J, Kircher M, et al. (Февраль 2016). "Ancient gene flow from early modern humans into Eastern Neanderthals". Природа. 530 (7591): 429–33. Bibcode:2016Natur.530..429K. Дои:10.1038/nature16544. ЧВК 4933530. PMID 26886800.
- Liu H, Prugnolle F, Manica A, Balloux F (August 2006). "A geographically explicit genetic model of worldwide human-settlement history". Американский журнал генетики человека. 79 (2): 230–7. Дои:10.1086/505436. ЧВК 1559480. PMID 16826514.
- Liu W, Martinón-Torres M, Cai YJ, Xing S, Tong HW, Pei SW, et al. (Октябрь 2015 г.). "The earliest unequivocally modern humans in southern China" (PDF). Природа. 526 (7575): 696–9. Bibcode:2015Natur.526..696L. Дои:10.1038/nature15696. PMID 26466566.
- Macaulay V, Hill C, Achilli A, Rengo C, Clarke D, Meehan W, et al. (May 2005). "Single, rapid coastal settlement of Asia revealed by analysis of complete mitochondrial genomes" (PDF). Наука. 308 (5724): 1034–6. Bibcode:2005Sci...308.1034M. Дои:10.1126/science.1109792. PMID 15890885.
- McChesney KY (2015). "Teaching Diversity. The Science You Need to Know to Explain Why Race Is Not Biological". SAGE Открыть. 5 (4). Дои:10.1177/2158244015611712.
- Meredith M (2011). Born in Africa: The Quest for the Origins of Human Life. New York City: PublicAffairs. ISBN 978-1-58648-663-1.
- Posth C, Renaud G, Mittnik A, Drucker DG, Rougier H, Cupillard C, et al. (Март 2016 г.). "Pleistocene Mitochondrial Genomes Suggest a Single Major Dispersal of Non-Africans and a Late Glacial Population Turnover in Europe". Текущая биология. 26 (6): 827–33. Дои:10.1016/j.cub.2016.01.037. HDL:2440/114930. PMID 26853362.
- Shackelton LA, Rambaut A, Pybus OG, Holmes EC (October 2006). "JC virus evolution and its association with human populations". Журнал вирусологии. 80 (20): 9928–33. Дои:10.1128/JVI.00441-06. ЧВК 1617318. PMID 17005670.
- Shen G, Wang W, Wang Q, Zhao J, Collerson K, Zhou C, Tobias PV (December 2002). «U-серия датировки сайта гоминидов Люцзян в Гуанси, Южный Китай». Журнал эволюции человека. 43 (6): 817–29. Дои:10.1006 / jhev.2002.0601. PMID 12473485.
- Wells S (2003) [2002]. The Journey of Man: A Genetic Odyssey. New York: Random House Trade Paperbacks. ISBN 978-0-8129-7146-0.
дальнейшее чтение
- Chris S (2011). The Origin of Our Species. Лондон: Аллен Лейн. ISBN 978-1-84614-140-9.
- Wells S (2006). Deep ancestry: inside the Genographic Project. Washington, D.C: National Geographic. ISBN 978-0-7922-6215-2.
- Wade N (2006). Before the Dawn : Recovering the Lost History of Our Ancestors. Penguin Press HC, The. ISBN 978-1-59420-079-3.
- Sykes B (2004). Семь дочерей Евы: наука, раскрывающая наше генетическое происхождение. Corgi Adult. ISBN 978-0-552-15218-1.
внешняя ссылка
- Британская энциклопедия, Эволюция человека
- Human Timeline (Interactive) – Смитсоновский институт, Национальный музей естественной истории (Август 2016 г.).
| Таксономия (Hominins ) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Предки |
| ||||||||||||||||||||||||||
| Модели |
| ||||||||||||||||||||||||||
| Сроки | |||||||||||||||||||||||||||
| Другие |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| Подтемы | |
|---|---|
| Генетическая история по регионам | |
| Популяционная генетика по группе |
|
| |