Homo ergaster - Homo ergaster
| Homo ergaster | |
|---|---|
 | |
| KNM-ER 3733, череп 1,6 миллиона лет назад Homo ergaster обнаружен в 1975 г. на Кооби Фора, Кения | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Haplorhini |
| Инфразаказ: | Симиобразные |
| Семья: | Гоминиды |
| Подсемейство: | Гоминины |
| Племя: | Гоминини |
| Род: | Гомо |
| Разновидность: | †Х. эргастер |
| Биномиальное имя | |
| †Homo ergaster | |
| Синонимы | |
† Telanthropus capensis[1] | |
Homo ergaster вымерший разновидность или же подвид из архаичные люди кто жил в Африке в Ранний плейстоцен. Ли Х. эргастер представляет собой отдельный вид или должен быть отнесен к H. erectus это продолжающийся и нерешенный спор в палеоантропология. Сторонники синонимизации обычно обозначают Х. эргастер в качестве "Африканский человек прямоходящий"[2] или же "Человек прямоходящий ergaster".[3] Название Homo ergaster примерно переводится как "работающий человек ", ссылка на более совершенные инструменты, используемые видом по сравнению с инструментами их предков.
Ареал ископаемых Х. эргастер в основном охватывает период от 1,8 до 1,7 миллиона лет назад, с несколькими более старыми и более молодыми экземплярами, расширяющими диапазон примерно до 2 и 1,5 миллиона лет назад соответственно. Хотя окаменелости известны по всей Восточной и Южной Африке, большинство Х. эргастер окаменелости были найдены на берегах Озеро Туркана в Кении. Есть более поздние африканские окаменелости, некоторые моложе 1 миллиона лет назад, которые указывают на долгосрочную анатомическую преемственность, хотя неясно, можно ли их формально рассматривать как Х. эргастер образцы. Как хроновиды, Х. эргастер возможно, сохранились до 600000 лет назад, когда новые родословные Гомо возник в Африке.
Те, кто верит Х. эргастер следует отнести к H. erectus считают, что между ними слишком мало различий, чтобы разделить их на отдельные виды. Сторонники сохранения двух видов в отдельности ссылаются на морфологические различия между африканскими окаменелостями и H. erectus окаменелости из Азии, а также ранние Гомо эволюция более сложна, чем то, что подразумевается при включении таких видов, как Х. эргастер в H. erectus. Кроме того, морфологические различия между образцами, обычно рассматриваемые как составляющие Х. эргастер мог бы предположить, что Х. эргастер сам по себе не представляет собой сплоченный вид. Независимо от их наиболее правильной классификации, Х. эргастер проявлять примитивные версии черт, позже выраженных в H. erectus и поэтому, вероятно, являются прямыми предками более поздних H. erectus популяции в Азии. Кроме того, Х. эргастер вероятно, является наследником более позднего гоминины в Европе и Африке, например современные люди и Неандертальцы.
Различают несколько особенностей Х. эргастер из австралопитеки а также более ранние и более базальные виды Гомо, Такие как H. habilis. Среди этих особенностей - большая масса тела, относительно длинные ноги, облигатные формы. двуногие относительно небольшие челюсти и зубы (что указывает на серьезное изменение в диете), а также пропорции тела и предполагаемый образ жизни, более похожий на современных людей, чем на более ранних и современных гомининов. Имея в виду эти особенности, некоторые исследователи считают, что Х. эргастер как самый ранний истинный представитель рода Гомо.
Х. эргастер жили в саванне в Африке, уникальной среде с проблемами, которые привели бы к необходимости многих новых и отличных форм поведения. Ранее Гомо вероятно, использовал тактику контратаки, как современные приматы, чтобы отпугнуть хищников. Ко времени Х. эргастер, такое поведение, вероятно, привело к развитию истинного охотник-собиратель поведение, первое среди приматов. Дальнейшее поведение, которое могло впервые возникнуть в Х. эргастер включать мужские и женские подразделения кормодобывания и истинные моногамный парные облигации. Х. эргастер также отмечает появление более совершенных инструментов Ашельский промышленность, включая самые ранние известные ручные топоры. Хотя неоспоримые доказательства отсутствуют, Х. эргастер также мог быть первым гоминином, мастер управления огнем.
Таксономия
История исследований

Систематика и систематика Гомо в раннем и среднем Плейстоцен является одним из самых спорных направлений палеоантропология.[4] В ранней палеоантропологии и вплоть до двадцатого века обычно считалось, что Х. сапиенс был конечным результатом постепенных модификаций внутри одной линии эволюции гомининов. Как воспринимаемая переходная форма между ранними гомининами и современными людьми, H. erectus, первоначально возведенный для хранения древних человеческих окаменелостей в Азии, стал охватывать широкий спектр окаменелостей, охватывающих большой промежуток времени (почти весь временной диапазон Гомо). С конца двадцатого века разнообразие внутри H. erectus заставил некоторых задуматься, что именно определяет вид и что он должен включать. Некоторые исследователи, например палеоантрополог Ян Таттерсолл в 2013 году задали вопросы H. erectus поскольку он содержит «громоздкое» количество окаменелостей с «существенно различающейся морфологией».[5]
В 1970-х годах палеоантропологи Ричард Лики и Алан Уокер описал серию окаменелостей гомининов из кенийских окаменелостей на восточном берегу Озеро Туркана. Наиболее заметными находками были два частичных черепа; KNM ER 3733 и KNM ER 3883, найдено в Кооби Фора. Лики и Уокер назначили эти черепа H. erectus, отмечая, что их объемы мозга (848 и 803 куб. см соответственно) хорошо сравниваются с образцом гораздо более молодого типа. H. erectus (950 куб. См). Еще одной важной окаменелостью была окаменелость нижняя челюсть восстановлен в Илерет и описан Лики с обозначением KNM ER 992 в 1972 г. как "Гомо неопределенных видов ».[6]
В 1975 году палеоантропологи Колин Гроувс и Вратислав Мазак обозначил KNM ER 992 как голотип образец особого вида, которого они назвали Homo ergaster.[7] Название (эргастер происходит из Древнегреческий ἐργαστήρ, Ergastḗr, «рабочий») примерно переводится как «рабочий»[8] или "рабочий".[9] Гровс и Мазак также включили многие окаменелости Фора Кооби, такие как KNM ER 803 (частичный скелет и некоторые изолированные зубы), в свое обозначение вида, но не предоставили никакого сравнения с азиатской летописью окаменелостей H. erectus в их диагнозе, непреднамеренно вызвав некоторую более позднюю таксономическую путаницу в отношении этого вида.[10]
Почти полная окаменелость, интерпретируемая как молодой мужчина (хотя пол на самом деле не определен), была обнаружена на западном берегу озера Туркана в 1984 году кенийским археологом. Камоя Кимеу.[9] Окаменелости были описаны Лики и Уокером вместе с палеантропологами Фрэнком Брауном и Джоном Харрисом в 1985 году как KNM-WT 15000 (по прозвищу «Мальчик Туркана»). Они истолковали окаменелость, состоящую из почти полного скелета, как представляющую H. erectus.[11] Мальчик Туркана был первым обнаруженным полностью сохранившимся экземпляром Х. эргастер/прямоходящий найдены и представляют собой важную окаменелость в установлении различий и сходств между ранними Гомо и современные люди.[12] Мальчик Туркана был помещен в Х. эргастер палеантропологом Бернардом Вудом в 1992 г.,[9] и сегодня наряду с другими окаменелостями в Африке ранее обозначался как H. erectus, обычно рассматривается как представитель Х. эргастер теми, кто поддерживает Х. эргастер как отдельный вид.[13]
Классификация
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Кладограмма на пролив, Грин и Флигл (2015)[14] |
Х. эргастер легко отличить от более ранних и более базальных видов Гомо, особенно H. habilis и Х. рудольфенсис по ряду черт, которые их объединяют, и предполагаемому образу жизни, более близко к современным людям, чем к более ранним и современным гомининам. По сравнению со своими родственниками, Х. эргастер имел пропорции тела, более похожие на более поздних представителей рода Гомо, особенно относительно длинные ноги, которые сделали бы их обязательно двуногими. Зубы и челюсти Х. эргастер также относительно меньше, чем у H. habilis и Х. рудольфенсис, что указывает на серьезные изменения в диете.[15] В 1999 году палеоантропологи Бернард Вуд и Марк Коллард утверждали, что общепринятые критерии отнесения видов к роду Гомо были ошибочными, и эти ранние и базальные виды, такие как H. habilis и Х. рудольфенсис, может быть надлежащим образом реклассифицирован как наследственный австралопитеки. По их мнению, самый ранний истинный представитель Гомо был Х. эргастер.[16]


С момента его описания как отдельного вида в 1975 году классификация окаменелостей Х. эргастер оспаривается. Х. эргастер был немедленно уволен Лики и Уокером и многими влиятельными исследователями, такими как палеоантрополог Дж. Филип Райтмайр, который написал обширный трактат по H. erectus в 1990 г. продолжал отдавать предпочтение более инклюзивной и всеобъемлющей H. erectus. В целом нет сомнений в том, что группа окаменелостей, составляющих H. erectus и Х. эргастер представляют собой окаменелости более или менее сплоченной группы близкородственных архаичных людей. Вместо этого возникает вопрос, представляют ли эти окаменелости излучение разных видов или излучение одного, очень изменчивого и разнообразного вида на протяжении почти двух миллионов лет.[7] Эта давняя дискуссия остается нерешенной, и исследователи обычно используют термины H. erectus SS. (Sensu stricto ) ссылаясь на H. erectus окаменелости в Азии и термин H. erectus s.l. (Sensu lato ) для обозначения окаменелостей других видов, которые могут или не могут быть включены в H. erectus, Такие как Х. эргастер, H. предшественник и Х. heidelbergensis.[17]
По понятным причинам Х. эргастер имеет много общего с H. erectus, такие как большие выступающие вперед челюсти, большие надбровные дуги и покатый лоб.[18] Многие особенности Х. эргастер явно более примитивные версии функций, позже выраженных в H. erectus, что несколько скрывает различия между ними.[19] Между окаменелостями Восточной Африки и Восточной Азии есть тонкие, потенциально значимые различия. Среди них черепа с несколько более высокими куполами и более тонкими стенками. Х. эргастер, а еще более массивный надбровные дуги и лица азиатских H. erectus.[18]
Вопрос усложняется, поскольку он касается того, сколько внутривидовых вариаций может проявиться у одного вида, прежде чем его нужно будет разделить на другие, вопрос, который сам по себе не имеет однозначного ответа. В 2008 году антрополог Карен Л. Бааб провела анализ окаменелостей различных H. erectus подвидов, включая окаменелости, приписываемые Х. эргастер, обнаружили, что внутривидовая изменчивость в пределах H. erectus было больше, чем ожидалось для одного вида по сравнению с современными люди и шимпанзе, но вполне укладывались в ожидаемые для вида вариации по сравнению с гориллы, и даже в пределах ожидаемого для одного подвида диапазона по сравнению с орангутаны (хотя отчасти это связано с большим половой диморфизм выставляется на горилл и орангутангов).[20] Бааб пришел к выводу, что H. erectus s.l. был либо единственным, но изменчивым видом, несколькими подвидами, разделенными по времени и географическому положению, либо несколькими географически рассредоточенными, но близкородственными видами.[21] В 2015 году палеантропологи Дэвид Стрейт, Фредерик Грин и Джон Флигл перечислили Х. эргастер как один из семи «широко известных» видов Гомо, рядом с H. habilis, Х. рудольфенсис, H. erectus, Х. heidelbergensis, H. neanderthalensis и Х. сапиенс, отмечая, что другие виды, такие как H. floresiensis и H. предшественник, были менее широко признаны или менее известны.[14]
Изменения в ископаемом материале



Сравнение различных африканских окаменелостей, приписываемых H. erectus или же Х. эргастер к азиатским окаменелостям, особенно типичный образец H. erectus, в 2013 году Ян Таттерсолл пришел к выводу, что, говоря об африканском материале как Х. эргастер а не "африканский H. erectus"было" значительным улучшением ", поскольку было много аутапоморфий, отличающих материал двух континентов друг от друга.[22] Таттерсолл считает целесообразным использовать обозначение H. erectus только для восточноазиатских окаменелостей, не обращая внимания на его предыдущее использование в качестве названия адаптивного сорта человеческих окаменелостей со всей Африки и Евразии. Хотя Таттерсолл пришел к выводу, что Х. эргастер материал представляет собой окаменелости одной клады Гомоон также обнаружил, что внутри этой клады имеется значительное разнообразие; нижняя челюсть KNM ER 992 хорошо сочетается с другими ископаемыми нижними челюстями из этого региона, такими как ОН 22 из Олдувай и KNM ER 3724 из Кооби Фора, но не обязательно совпадал с черепным материалом, таким как KNM ER 3733 и KNM ER 3883 (так как не сохраняет челюсть), ни с сохранившейся нижней челюстью у Turkana Boy, у которого заметно отличается зубной ряд.[22]
Самая «культовая» окаменелость Х. эргастер череп KNM ER 3733, резко отличающийся от азиатского H. erectus по ряду характеристик, включая то, что надбровные дуги выступают вперед и вверх и дуги отдельно по каждой орбите, а черепная коробка довольно высока по сравнению с ее шириной, а ее боковые стенки изогнуты. KNM ER 3733 можно отличить от KNM ER 3883 по ряду особенностей, в частности, в том, что края надбровных дуг KNM ER 3883 очень утолщены и выступают наружу, но слегка вниз, а не вверх.[23] Оба черепа можно отличить от черепа Turkana Boy, который имеет лишь незначительно значительные утолщения верхних краев глазницы, без более вертикального утолщения KNM ER 3883 и агрессивного выступа KNM ER 3733. В дополнение к этому, структура лица Мальчика Туркана уже и длиннее, чем у других черепов, с более высоким носовым отверстием и, вероятно, более плоским профилем верхней части лица. Возможно, что эти различия можно объяснить тем, что мальчик Туркана был несовершеннолетним в возрасте от 7 до 12 лет.[24] Кроме того, предполагается, что KNM ER 3733 был черепом женщины (в то время как мальчик Туркана традиционно интерпретируется как мужчина), что означает, что половой диморфизм может объяснять некоторые различия.[12]
Различия между черепом Turkana Boy и KNM ER 3733 и KNM ER 3883, а также различия в зубных рядах между Turkana Boy и KNM ER 992 были интерпретированы некоторыми, такими как палеантрополог Джеффри Х. Шварц, как предполагающие, что Turkana Boy и Остальные Х. эргастер материал не представляет один и тот же таксон. Шварц также отметил, что ни одна из окаменелостей не представляет H. erectus либо, который, по его мнению, нуждался в значительном пересмотре.[25] В 2000 году французский палеоантрополог Валери Зейтун предложила отнести KNM ER 3733 и KNM ER 3883 к двум отдельным видам, которые она назвала H. kenyanensis (типовой образец КНМ ЭР 3733) и H. okotensis (типовой образец KNM ER 3883), но эти обозначения не нашли широкого распространения.[26]
Эволюционная история
Эволюция и временной диапазон
Хотя часто предполагается, что они возникли в Восточная Африка, истоки Х. эргастер затушевываются тем фактом, что этот вид знаменует собой радикальное отличие от более ранних видов Гомо и Австралопитек в его длинных конечностях, росте и современных пропорциях тела. Хотя большое количество орудий плейстоцена было найдено в Восточной Африке, нельзя полностью установить, что Х. эргастер возник там без дальнейших открытий окаменелостей.[27] Предполагается, что Х. эргастер произошли от более ранних видов Гомо, наверное H. habilis. Хотя популяции Х. эргастер за пределами Африки были выведены на основе географического распределения их потомков и орудий, соответствующих таковым в Восточной Африке, окаменелости этого вида в основном происходят из Восточной Африки в диапазоне времени от 1,8 до 1,7 миллиона лет назад. Большинство окаменелостей было обнаружено у берегов озера Туркана в Кении.[8]
Самый старый известный образец H. erectus s.l. в Африке (т.е. Х. эргастер) является DNH 134, череп, найденный в Дримолен Система Палеокавы в Южной Африке, датированная более 1,95 миллиона лет назад. Череп также является старейшим из известных H. erectus s.l. образец в целом, демонстрирующий явное сходство с KNM ER 3733 и демонстрирующий, что Х. эргастер сосуществовали с другими гомининами, такими как Paranthropus robustus и Австралопитек седиба.[28]
Встречаются и более молодые экземпляры Х. эргастер; примечательно, что возраст Turkana Boy составляет около 1,56 миллиона лет назад.[8] Горстка африканских черепов еще более молодого возраста свидетельствует о долгосрочной анатомической преемственности, хотя неясно, можно ли их формально рассматривать как Х. эргастер образцы; "Олдувайский гоминид 9 «череп из Олдувайского ущелья датируется примерно 1,2–1,1 миллиона лет назад, и есть также черепа из Буйи (около побережья Эритреи, датируемые примерно 1 миллион лет), формации Бури в Эфиопии (датируемой от 1 миллиона до 780 000 лет) и фрагмент черепа из Олоргесайли в Кении (датированный 970 000–900 000 лет назад). Олдувайский череп похож на азиатский. H. erectus в его массивной надбровной дуге, но остальные лишь незначительно отличаются от более ранних Х. эргастер черепа.[29]
В H. erectus в Азии, а также более поздние гоминины в Европе (т.е. Х. heidelbergensis и H. neanderthalensis) и Африке (Х. сапиенс) все, вероятно, произошли от Х. эргастер.[8] Потому что Х. эргастер считается предком этих более поздних Гомо, он мог сохраняться в Африке примерно до 600000 лет назад, когда размер мозга быстро увеличился и Х. heidelbergensis появился.[30]
Выход из Африки

Традиционно H. erectus считался гоминином, который первым покинул Африку, чтобы колонизировать Европу и Азию. Если Х. эргастер отличается от H. erectus, эта роль будет применяться к Х. эргастер вместо.[22][8] Известно очень мало конкретной информации о том, когда и какие Гомо впервые появились в Европе и Азии, поскольку ископаемые гоминины раннего плейстоцена редки на обоих континентах, и что это могло быть Х. эргастер (или "рано H. erectus"), который расширился, а также конкретный способ, которым они были расширены, остается предположением.[18] Наличие H. erectus окаменелости в Восточной Азии означает, что человеческий вид, скорее всего, Х. Эргастерпокинул Африку до 1 миллиона лет назад,[31] Исторически предполагалось, что они впервые мигрировали из Африки примерно от 1,9 до 1,7 миллиона лет назад.[18] Открытия в Грузии и Китае отодвигают самую последнюю возможную дату на 2 миллиона лет назад, что также ставит под сомнение идею о том, что Х. эргастер был первым гоминином, покинувшим Африку.[32]
Основная причина ухода из Африки, вероятно, заключалась в том, что рост населения, периодически превышающий их ресурсную базу, с отколовшимися группами, перемещающимися со временем, чтобы обосноваться на соседних, пустых территориях. Физиология и улучшенная технология Х. эргастер мог позволить им путешествовать и колонизировать территории, которые никто никогда раньше не занимал.[31] Неясно, если Х. эргастер была действительно уникальной способной к расширению за пределами Африки; австралопитеки, вероятно, колонизировали луга саванны по всей Африке 3 миллиона лет назад, и нет четких причин, почему они не смогли бы расшириться на луга Азии раньше Х. эргастер.[17]
Общее предположение состоит в том, что гоминины мигрировали с континента через южную оконечность красное море или вдоль Долина Нила, но нет никаких ископаемых гомининов, известных ни в одном из регионов в раннем плейстоцене. Раннее Гомо окаменелости за пределами Африки являются Черепа Дманиси из Грузии (возраст 1,77–1,85 млн лет,[32] представляющий либо ранний Х. эргастер или новый таксон, Х. georgicus ), три резца из Убейдия в Израиле (от 1,4 до 1 миллиона лет) и окаменелости Ява человек (H. erectus прямоходящий, более пяти тысяч миль).[27] Знакомства ключевых азиатских H. erectus количество экземпляров (включая Java Man) не совсем достоверно, но все они, вероятно, возрастом 1,5 миллиона лет или моложе.[18] Убейдия также является старейшим официально подтвержденным местом Ашельский инструменты (одна из инструментальных отраслей, связанных с Х. эргастер) за пределами Африки найденные там инструменты очень напоминали старые инструменты, обнаруженные в Восточной Африке.[31]
Самые ранние ископаемые свидетельства Гомо в Азии находятся вышеупомянутые черепа Дманиси, которые имеют много общих черт с Х. эргастер в Африке, предполагая, что Х. эргастер мог распространиться за пределы Африки уже 1,7–1,9 миллиона лет назад.[33] В добавление к Х. эргастер-подобные черты, черепа Дманиси обладают широким набором других черт, некоторые из которых похожи на черты более ранних гомининов, таких как H. habilis, а также отсутствие сохранившихся ручных топоров (в остальном характерных для Х. эргастер), что означает, что гоминины могли распространиться из Африки даже раньше, чем Х. эргастер.[17] Череп D2700 (Череп Дманиси 3) в особенности напоминает H. habilis в небольшом объеме его черепной коробки (600 куб. см), форме средней и верхней части лица и отсутствии внешнего носа. Смешение черепов в Дманиси предполагает, что определение Х. эргастер (или же H. erectus) наиболее целесообразно расширить, чтобы включить ископаемые, которые в противном случае были бы отнесены к H. habilis или что два отдельных вида архаичных людей рано покинули Африку.[29] Помимо окаменелостей Дманиси, на острове были обнаружены каменные орудия труда, изготовленные гомининами. Лессовое плато в Китае и датируется 2,12 миллионами лет, а это означает, что гоминины должны были покинуть Африку раньше этого времени.[32]
Исторически сложилось так, что альтернативная гипотеза Гомо развился в Азии от более ранних предков, которые мигрировали туда из Африки, а затем распространились обратно в Европу, где породили Х. сапиенс. Этого мнения особенно придерживались Эжен Дюбуа, кто первым описал H. erectus окаменелости в 19 веке и считались окаменелостями яванского человека, в то время несомненно самыми ранними из известных окаменелостей гомининов, в качестве доказательства гипотезы. Хотя открытие австралопитеков и ранее Гомо в Африке означало, что Гомо сама по себе не возникла в Азии, идея, что H. erectus (или же Х. эргастер), в частности, сделал, а затем расширился обратно в Африку, время от времени всплывал.[34] На протяжении многих лет для подтверждения этого использовались различные находки окаменелостей, возможно, самая известная из них - массивные челюсти из Индонезии, которые были сочтены похожими на челюсти австралопитеков и названы Мегантроп (теперь считается, что это неродственная обезьяна-гоминид). Открытие H. floresiensis в 2003 году, сохранивший примитивную анатомию стопы и запястья, напоминающую анатомию H. habilis и Австралопитек снова привел к предложениям пред-прямоходящий гомининов в Азии, хотя сопоставимых костей стопы или запястья из H. erectus что делает невозможным сравнение.[35] Идея, что Х. эргастер/H. erectus впервые возник в Азии до того, как вернуться в Африку, был существенно ослаблен датировкой черепа DNH 134 как приблизительно 2 миллиона лет, до всех других известных Х. эргастер/H. erectus окаменелости.[28]
Анатомия
Телосложение и внешний вид

Единственные хорошо сохранившиеся посткраниальные остатки Х. эргастер происходят из окаменелости мальчика Туркана.В отличие от австралопитеков, руки мальчика Туркана не были длиннее по сравнению с его ногами, чем руки живых людей, а конусообразное туловище его / ее предков превратилось в более бочкообразную грудь над узкими бедрами, что является еще одним сходством с современными. люди.[36] В большеберцовая кость (большеберцовая кость) мальчика Туркана относительно длиннее, чем такая же кость у современного человека, что потенциально означает, что при ходьбе колено сгибалось сильнее.[37] Стройное и длинное тело Turkana Boy можно объяснить: Х. эргастер живущие в жарких и засушливых сезонных условиях. Из-за истончения тела объем тела уменьшается быстрее, чем площадь кожи, а большая площадь кожи означает более эффективное рассеивание тепла.[38]
Х. эргастер люди были значительно выше своих предков. В то время как Люси, знаменитый Австралопитек окаменелости, на момент ее смерти был бы всего около 1 м (3 фута 3 дюйма) в высоту, Туркана Бой был около 1,62 м (5 футов 4 дюйма) в высоту и, вероятно, достиг бы 1,82 м (6 футов) или более, если бы он / она дожил до совершеннолетия.[36] Взрослый Х. эргастер считается, что их размер варьировался от 1,45 до 1,85 м (от 4 футов 9 дюймов до 6 футов 1 дюймов) в высоту.[37]
Благодаря адаптации к жаркому и засушливому климату, Х. эргастер Возможно, это был самый ранний человеческий вид, у которого была почти голая кожа. Х. эргастер имел обезьяноподобный покров волос на теле, потливость (основные средства, с помощью которых современные люди предотвращают перегрев своего мозга и тела) были бы не столь эффективны.[38] Хотя потоотделение является общепринятым объяснением отсутствия волос, другие предлагаемые объяснения включают уменьшение паразит нагрузка[39] и половой отбор.[40] Сомнительно, чтобы австралопитеки и ранее Гомо были достаточно подвижны, чтобы сделать выпадение волос выгодным признаком, тогда как Х. эргастер были явно адаптированы для дальних путешествий и были известны тем, что населяли более низкие высоты (и открытые, жаркие среды саванн), чем их предки. Австралопитеки обычно населяли более холодные и более высокие высоты 1 000–1600 м (3 300–5 200 футов), где ночные температуры могли бы стать значительно ниже и, возможно, потребовались бы изолирующие волосы на теле.[41]
В противном случае, несмотря на это, выпадение волос на теле могло произойти значительно раньше, чем Х. эргастер. Хотя отпечатки кожи неизвестны ни у одного вымершего гоминина, вполне возможно, что предки человека уже теряли волосы на теле около 3 миллионов лет назад. Приобретены предки человека лобковые вши от горилл около 3 миллионов лет назад, а видообразование человека от лобковых вшей гориллы было потенциально возможно только потому, что предки человека к тому времени потеряли большую часть волос на теле.[42] Также возможно, что выпадение волос на теле произошло значительно позже. Генетический анализ показывает, что высокая активность в рецептор меланокортина 1, дающий темную кожу, появился примерно 1,2 миллиона лет назад. Это может указывать на эволюцию безволосости примерно в это время, так как отсутствие волос на теле оставило бы кожу подверженной вредному воздействию. УФ-излучение.[43]
Череп и лицо

Отличия от современных людей были бы очевидны по лицу и черепу Х. эргастер. К моменту его / ее смерти мозг мальчика Туркана был почти полностью развит, но его объем (880 куб. См) был всего на 130 куб. H. habilis, примерно на 500 куб. см ниже среднего уровня современного человека. Увеличение на 130 куб. См по сравнению с H. habilis становится гораздо менее значительным, чем то, что можно было бы предположить, когда больший размер тела мальчика Туркана и Х. эргастер Считается.[44] Со всем Х. эргастер Если рассматривать черепа, то объем мозга у этих видов в основном варьировался от 600 до 910 куб. см, а некоторые небольшие образцы имели объем только 508–580 куб. Поскольку их мозг был меньше, чем у современных людей, череп Х. эргастер сразу зауженные за глазницами (посторбитальное сужение ).[37]
Головной мозг был длинным и низким, а лоб мальчика Туркана был плоским и скошенным, сливаясь под углом с надбровной дугой над его / ее глазами. Заметная разница между Турканским мальчиком и австралопитеками и H. habilis это был бы его / ее нос, который был бы похож на нос современных людей в том, что он выглядел бы вперед и имел бы ноздри, ориентированные вниз. Этот внешний нос, возможно, также был адаптацией к более теплому климату, поскольку носы современных людей обычно холоднее, чем их центральные тела, и конденсирует влагу, которая в противном случае выдыхалась бы и терялась в периоды повышенной активности.[44]Лицо мальчика Туркана было бы длиннее сверху вниз, чем у современного человека, а челюсти выступали бы наружу (прогнатизм ). Хотя челюсти и зубы были меньше, чем у среднего австралопитека и H. habilis, они все еще были значительно больше, чем у современного человека. Поскольку челюсть резко отклонена назад, вероятно, у него не было подбородка.[38]
Общая структура черепа и лица мальчика Туркана также отражена в других Х. эргастер черепа, в которых большие и выступающие наружу лица сочетаются с надбровными дугами, скошенными лбами, большими зубами и выступающими носовыми костями.[38] Хотя мальчику Туркана на момент смерти было бы не более 12 лет, его рост больше похож на рост современного 15-летнего ребенка, а мозг сопоставим с мозгом современного годовалого ребенка. . По современным меркам, Х. эргастер таким образом, были бы когнитивно ограничены, хотя изобретение новых инструментов доказывает, что они были умнее своих предшественников.[45]
Масса тела и половой диморфизм

Х. эргастер обладали значительно большей массой тела по сравнению с более ранними гомининами, такими как ранние Гомо, Австралопитек и Парантроп.[15] В то время как австралопитеки обычно имели вес от 29 до 48 кг (от 64 до 106 фунтов), Х. эргастер обычно весил 52–63 кг (115–139 фунтов).[46] Возможно, что увеличенный размер тела был результатом жизни в открытой среде саванны, где увеличенный размер дает возможность использовать более широкий рацион в более крупных районах кормления, увеличивает подвижность, а также дает возможность охотиться на более крупную добычу.[15] Увеличенная масса тела также означает, что родители смогли бы отнести своих детей к более старшему возрасту и большей массе.[46]
Хотя снижение полового диморфизма исторически часто упоминалось как одно из радикальных различий между Х. эргастер и раньше Гомо и австралопитеки,[15][27] неясно, были ли австралопитеки значительно более сексуально диаморфными, чем Х. эргастер или современные люди.[47] Скелетные данные свидетельствуют о том, что пол в Х. эргастер различались по размеру не больше, чем пол современного человека,[45] но исследование 2003 года, проведенное палеоантропологами Филипом Л. Рино, Ричардом С. Майндлом, Мелани А. Макколлум и Ч. Оуэном Лавджоем, показало, что то же самое верно и для значительно более ранних Австралопитек афарский.[47] Половой диморфизм у вымерших видов трудно измерить, поскольку пол ископаемых обычно не поддается определению. Исторически сложилось так, что ученые обычно измеряли различия между крайними концами (с точки зрения размера и морфологии) ископаемого материала, приписываемого виду, и предполагали, что полученное соотношение относится к средней разнице между мужчинами и женщинами.[48]
Рост и развитие
Размеры взрослой женщины возрастом 1,8 миллиона лет Х. эргастер таз от Гона, Эфиопия предполагает, что Х. эргастер были способны родить детей с максимальным пренатальным (до рождения) размером мозга 315 куб. см, что составляет около 30–50% от размера мозга взрослого человека. Это значение находится где-то между шимпанзе (~ 40%) и современным человеком (28%).[49] Дальнейшие выводы о росте и развитии в начале Гомо можно извлечь из Mojokerto ребенок, ~ 1,4–1,5 миллиона лет ~ годовалый азиат H. erectus, мозг которого составлял 72–84% от размера взрослого H. erectus мозг, что предполагает траекторию роста мозга, более похожую на траекторию роста других большие обезьяны чем у современных людей.[50] И таз Гона, и ребенок Моджокерто предполагают, что пренатальный рост из Х. эргастер был подобен таковому у современных людей, но послеродовой (послеродовой) рост и развитие занимали промежуточное положение между шимпанзе и современными людьми.[49] Более высокая скорость развития предполагает, что альтрициальность (продолжительное детство и длительный период зависимости от родителей) развились на более позднем этапе эволюции человека, возможно, у последнего общего предка неандертальцев и современных людей.[50] Более высокая скорость разработки может также указывать на то, что ожидаемая продолжительность жизни Х. эргастер и H. erectus был ниже, чем у более поздних и современных людей.[51]
Культура
Диета и энергетика
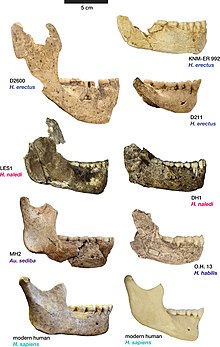
Часто предполагается, что больший размер тела и мозга Х. эргастер, по сравнению с его предками, принесла бы с собой повышенные потребности в питании и энергии.[46] В 2002 году палеоантропологи Лесли К. Айелло и Джонатан К. К. Уэллс заявили, что средние метаболические потребности в покое Х. эргастер были бы на 39% выше, чем у Австралопитек афарский, На 30% выше у мужчин и на 54% выше у женщин.[52] Однако пропорции туловища Х. эргастер подразумевает относительно небольшой кишечник,[52] это означает, что потребности в энергии не обязательно были выше в Х. эргастер чем у более ранних гомининов. Это связано с тем, что кишечник более ранних обезьян (и австралопитеков) был большим и требовал больших затрат энергии, так как ему нужно было синтезировать жир путем ферментации растительного вещества, тогда как Х. эргастер вероятно, ели значительно больше животного жира, чем их предшественники. Это позволило бы направить больше энергии на рост мозга, увеличив размер мозга, сохранив при этом потребности в энергии более ранних видов.[53][54]
Если бы у них была повышенная потребность в энергии, Х. эргастер пришлось бы съесть либо гораздо больше пищи, чем австралопитеки, либо потребовалось бы есть пищу высшего качества. Если бы они ели такую же пищу, что и австралопитеки, время кормления пришлось бы резко увеличить пропорционально требуемым дополнительным калориям, сокращая время Х. эргастер можно использовать для отдыха, общения и путешествий. Хотя это было бы возможно, это считается маловероятным, тем более что челюсти и зубы Х. эргастер имеют меньший размер по сравнению с таковыми у австралопитеков, что предполагает отказ от волокнистой и трудно пережевываемой пищи. Независимо от потребностей в энергии, тонкая кишка Х. эргастер также предлагает более легко усваиваемую диету, состоящую из продуктов более высокого качества.[52]
Похоже, что Х. эргастер потребляли мясо в более высоких пропорциях, чем более ранние австралопитеки.[52] Мясо, вероятно, было добыто в результате сочетания засад, активной охоты и конфронтации с мусором. Х. эргастер должны были не только обладать способностью к выносливости бега, но также должны были уметь защищать себя и трупы своей добычи от множества современных африканских хищников. Не исключено, что капля африканского карниворан Видовое разнообразие около 1,5 миллионов лет назад можно отнести к конкуренции с оппортунистическими и хищными гомининами.[55]
Само по себе мясо могло не выдержать Х. эргастер. Современные люди не могут в достаточной степени усваивать белок, чтобы удовлетворить более 50% своих энергетических потребностей, а современные люди, которые в значительной степени полагаются на продукты животного происхождения в своем рационе, в основном полагаются на жиры для удовлетворения остальных своих энергетических потребностей.[52] Множество причин делают полностью мясную диету Х. эргастер маловероятно, что самым известным из них был африканский копытные (основная доступная добыча) имеют относительно низкое содержание жира, и что диета с высоким содержанием мяса требует повышенного потребления воды, что было бы трудно в открытой и жаркой среде. Современный африканский охотники-собиратели которые сильно полагаются на мясо, например Хадза и Сан народы, также используют культурные средства для извлечения максимального количества жира из туш своей добычи, метод, который был бы недоступен для Х. эргастер.[56]
Х. эргастер таким образом, вероятно, потребляли бы большое количество мяса, намного больше, чем их предки, но им также пришлось бы использовать множество других источников пищи, таких как семена, мед, орехи, беспозвоночные,[56] питательные клубни, луковицы и другие запасающие органы подземных растений.[38] Относительно небольшая жевательная способность Х. эргастер, по сравнению с его предками с большей челюстью, означает, что потребляемые мясо и высококачественная растительная пища, вероятно, потребовали бы использования инструментов для обработки перед едой.[57]
Социальная структура и динамика

Х. эргастер жили в африканской саванне, которая в плейстоцене была домом для гораздо более грозного сообщества хищников, чем нынешняя саванна. Гоминины, вероятно, могли бы адаптироваться к жизни в саванне только в том случае, если бы эффективная защита от хищников уже сформировалась. Защита от хищников, вероятно, прошла бы Х. эргастер жизнь в больших группах, владение каменными (и, предположительно, деревянными) инструментами и эффективное контратакующее поведение. У современных приматов, которые проводят значительное количество времени в саванне, например, у шимпанзе и саванна бабуины, особи образуют большие группы, состоящие из нескольких самцов, в которых несколько самцов могут эффективно работать вместе, чтобы отбиваться и контратаковать хищников, иногда с использованием камней или палочек, и защищать остальную часть группы. Возможно, подобное поведение проявлялось в раннем Гомо. Основываясь на системе привязанности к самцам у бонобо и шимпанзе и тенденции к привязке самцов у современных собирателей, группы ранних Гомо возможно, тоже были связаны с мужчиной. Из-за нехватки ископаемого материала размер группы в начале Гомо невозможно определить с какой-либо уверенностью. Группы, вероятно, были большими, возможно, группы были выше верхнего диапазона известных размеров групп среди шимпанзе и бабуинов (c. 100 человек и более).[58] В 1993 году палеоантропологи Лесли К. Айелло и Р. И. М. Данбар подсчитали, что размер группы H. habilis и Х. рудольфенсис, на основе неокортекс размер (поскольку существует известная взаимосвязь между размером неокортекса и размером группы у современных нечеловеческих приматов),[59] колеблется от 70 до 85 человек.[60] С дополнительным фактором двуногости, который энергетически дешевле, чем четвероногий, максимально приемлемый с экологической точки зрения размер группы мог быть даже больше.[58] Оценка размера группы Айелло и Данбара в отношении Х. эргастер 91–116 человек.[60]
Социальное и контр-атакующее поведение ранее Гомо вероятно перенесен в Х. эргастер, где они, вероятно, получили дальнейшее развитие. Х. эргастер был, вероятно, первым приматом, занявшим нишу социальных хищников (т. е. охотник-собиратель ).[58] Такое поведение, вероятно, было бы результатом контратак в контексте конкуренции за питательную пищу с другими плотоядными животными и, вероятно, эволюционировало бы из чего-то похожего на оппортунистическую охоту, которую иногда проявляют шимпанзе. Переход к групповому хищничеству мог спровоцировать каскад эволюционных изменений, которые изменили ход человеческой эволюции. Совместное поведение, такое как оппортунистическая охота в группах, защита хищников и конфронтационная уборка мусора, имели решающее значение для выживания, а это означает, что постепенно произошел фундаментальный переход в психологии. С типичным поведением «конкурентного сотрудничества», демонстрируемым большинством приматов, больше не поощряемым естественным отбором и сменой социальных тенденций, охота и другие виды деятельности превратились бы в настоящие совместные усилия. Поскольку у самцов современных приматов обычно проявляется контратакующее поведение, социальная охота у архаичных людей, как полагают, была преимущественно мужской деятельностью. Самки, вероятно, занимались другими видами кормодобывания, собирая пищу, не требующую охоты (например, фрукты, орехи, яйца и т. Д.).[61]
Поскольку охота была социальной деятельностью, люди, вероятно, делили мясо друг с другом, что укрепило бы связи как между самими охотниками, так и между охотниками и остальной частью населения. Х. эргастер группа. Самки, вероятно, делились тем, что добывали, с остальной группой. Такое развитие событий могло привести к превращению мужской и женской дружбы в оппортунистическую. моногамный парные облигации. Поскольку половой отбор от самок, вероятно, благоприятствовал самцам, которые могли охотиться, возникающее социальное поведение, возникающее в результате этих новых форм поведения, передавалось и усиливалось из поколения в поколение.[61]
Единственное прямое свидетельство Х. эргастер Состав группы происходит из ряда стоянок за пределами Илерета в Кении, где сохранилось 97 следов, оставленных около 1,5 миллиона лет назад группой из не менее 20 человек. Судя по размеру следов, одна из тропинок представляла собой группу, полностью состоящую из мужчин, возможно, специализированную рабочую группу, такую как пограничный патруль или группа охотников или собирателей. Если эта оценка верна, это также предполагает разделение обязанностей между мужчинами и женщинами. В современном охотник-собиратель В обществах, нацеленных на крупную добычу, группы мужчин обычно отправляются для уничтожения этих животных с высоким риском, а из-за низкого уровня успеха женские группы, как правило, сосредотачиваются на более предсказуемой пище.[62]
Технологии
Инструментальное производство

Рано Х. эргастер унаследовал Oldowan культура орудий труда австралопитеков и ранее Гомо, хотя быстро научились поражать каменные отщепы гораздо большего размера, чем их предшественники и современники. 1,65 миллиона лет назад Х. эргастер создали обширные чешуйчатые артефакты и ранние ручные топоры которые отмечают ашельскую культуру,[8] 1,6–1,4 миллиона лет назад новая инструментальная промышленность получила широкое распространение в Восточной Африке.[63] Ашельские инструменты отличаются от олдовских инструментов тем, что основные формы инструментов были явно преднамеренными. В то время как форма стержней в орудиях Олдува, которые, вероятно, использовались в основном в качестве молотков для раскалывания костей для костного мозга, похоже, не имела большого значения, ручные топоры ашельской культуры демонстрируют намерение производить узкие и острые предметы, обычно каплевидной, овальной или треугольной формы.[30] Оказавшись на месте, ашельская промышленность оставалась неизменной на протяжении всего периода Х. эргастерСуществование и более поздние времена, когда инструменты, произведенные около 250 000 лет назад, не сильно отличаются от инструментов, произведенных 1,65 миллиона лет назад.[64]

Самые древние ашельские комплексы также сохраняют основные формы, аналогичные тем, что используются в олдовских орудиях, но нет никаких известных истинных промежуточных форм между ними, что позволяет предположить, что появление ашельских орудий было резким и внезапным развитием. Наиболее значительным развитием, которое привело к созданию ашельских орудий, были, вероятно, первые гоминины, которые научились наносить удары по крупным чешуйкам длиной до 30 см (1 фут) или более из более крупных валунов, из которых они могли изготавливать новые инструменты, такие как ручные топоры. .[63] Хотя «ручной топор» подразумевает, что все ручные топоры использовались для рубки и были ручными, они были разных форм и размеров и, вероятно, выполняли несколько различных функций. Тщательно сформированные и симметричные образцы могли быть брошены в жертву, сродни современной обсуждает, более простые образцы могли просто служить переносными источниками острых хлопьев, а некоторые могли использоваться для соскабливания или рубки древесины. Кроме того, ручные топоры - эффективные инструменты разделки мяса и, возможно, также использовались для расчленения туш крупных животных.[64]
Сохранились ручные топоры, которые слишком громоздки и велики, чтобы их можно было использовать для каких-либо очевидных практических целей. Использование этих больших ручных топоров и некоторых обнаруженных коллекций из сотен ручных топоров без явных признаков использования является спекулятивным и предположительным. Идея, которая была популярна в популярной прессе и часто цитируется в академических кругах,[65] заключается в том, что большие и впечатляющие ручные топоры могли быть эмблемами, используемыми для привлечения товарищей, причем производители больших топоров демонстрировали силу, координацию и решительность - качества, которые могли считаться привлекательными.[64] Палеоантропологи Эйприл Новелл и Мелани Ли Чанг отметили в 2009 году, что, хотя эта теория «интригует и эмоционально привлекательна», в ее пользу мало свидетельств, и она не поддается проверке.[66] Они посчитали более вероятным, что изменения морфологии топора на протяжении сотен тысяч лет были результатом различных факторов, а не единственного всеобъемлющего фактора полового отбора.[67]
Огонь
В качестве Гомо мигрировали в открытые саванны, встречи с естественными пожарами должны были стать более частыми и значительными.[68] Возможно, что Х. эргастер был одним из первых людей, освоивших управление огнем, которое они, возможно, использовали для приготовления пищи. Приготовление делает мясную и растительную пищу более усвояемой, что, возможно, было важно, так как кишечник Х. эргастер были уменьшены в размерах по сравнению с таковыми их предков.[38] Хотя Х. эргастер/H. erectus часто считается, что это был самый ранний Гомо Для борьбы с огнем в летописи окаменелостей не хватает конкретных доказательств, возможно, отчасти из-за сложности сохранения фактических доказательств использования огня.[69][70] Двумя самыми ранними объектами, которые, как обычно утверждается, сохраняют свидетельства использования огня, являются FxJj20 на Куби Фора и GnJi 1 / 6E около Озеро Баринго как в Кении, так и возрастом до 1,5 миллионов лет. Свидетельства на FxJj20 состоят из обгоревших отложений и каменных орудий, подвергшихся тепловому изменению, тогда как GnJi 1 / 6E сохраняет большие Clasts из обожженной глины, связанные с каменными орудиями и остатками фауны. Хотя трудно исключить естественное происхождение обнаруженных остатков пожара, эти участки остаются сильными кандидатами на раннее использование пожаров.[71][70]
Несколько мест, сохраняющих более широко признанные свидетельства использования огня, были датированы 1 миллион лет назад или моложе, после появления и последней общепринятой записи Х. эргастер.[38] Эти сайты включают в себя пещеры, такие как Wonderwerk и Сварткранс в Южной Африке и на открытых сайтах, таких как Kalambo Falls в Замбии. На памятнике Гешер Бенот Яаков в Израиле, датируемом примерно 700 000 лет назад, сохранились широко признанные свидетельства использования огня в виде обожженных материалов и микрочастиц сгоревшего кремня, которые сохранились на многих уровнях.[71][38] Примерно с 400 000 лет назад и позже следов пожаров стало еще больше на территориях Африки, Европы и Азии.[72]
Язык
В спинной мозг мальчика Туркана было бы уже, чем у современного человека, а это означает, что нервная система Х. эргастери их дыхательные мышцы, возможно, не были достаточно развиты, чтобы производить или контролировать речь.[37] В 2001 году антропологи Брюс Латимер и Джеймс Оман пришли к выводу, что мальчик Туркана страдал от скелетная дисплазия и сколиоз, и, следовательно, не был бы представителем остальной части своего вида в этом отношении.[73] В 2006 году, когда антрополог Марк Мейер и коллеги описали H. erectus s.l. экземпляр из Дманиси, Грузия, возрастом 1,78 миллиона лет. В окаменелостях сохранились самые старые из известных Гомо Найденные позвонки и позвоночник попадают в диапазон современных человеческих позвоночников, что позволяет предположить, что этот человек был способен говорить. Мейер с коллегами пришли к выводу, что речь, вероятно, была возможна в Гомо очень рано, и этот мальчик Туркана, вероятно, страдал каким-то врожденным дефектом, возможно стеноз позвоночного канала.[74]
В 2013 и 2014 годах антрополог Регула Шисс и его коллеги пришли к выводу, что свидетельств каких-либо врожденные дефекты в Turkana Boy, и, в отличие от исследований 2001 и 2006 годов, считали, что этот экземпляр является представителем этого вида.[75][76]
Смотрите также
Рекомендации
- ^ Веник, Р.; Талбот, Дж. Т. (1949). «Новый тип ископаемого человека». Природа. 164 (4164): 322–323. Дои:10.1038 / 164322a0. PMID 18137042. S2CID 4106457.
- ^ Таттерсолл 2013, п. 5.
- ^ Таттерсолл 2013, п. 14.
- ^ Таттерсолл 2013, п. 1.
- ^ Таттерсолл 2013, п. 2.
- ^ Таттерсолл 2013, стр. 3–4.
- ^ а б Таттерсолл 2013, п. 4.
- ^ а б c d е ж Кляйн 2005, п. 85.
- ^ а б c Робертс 2018, п. 116.
- ^ Антон 2003, п. 127.
- ^ Brown et al. 1985 г., п. 788.
- ^ а б Brown et al. 1985 г., п. 789.
- ^ Wang et al. 2004 г., п. 453.
- ^ а б Пролив, Грин и Флигл 2015, п. 2006 г.
- ^ а б c d Айелло и Уэллс 2002, п. 324.
- ^ Дерево и коллард 1999, п. 65.
- ^ а б c Dennell & Roebroeks 2005, п. 1100.
- ^ а б c d е Кляйн 2005, п. 92.
- ^ Антон 2003, п. 154.
- ^ Бааб 2008, п. 841.
- ^ Бааб 2008, п. 842.
- ^ а б c Таттерсолл 2013, п. 15.
- ^ Таттерсолл 2013, п. 8.
- ^ Таттерсолл 2013, п. 9.
- ^ Шварц 2000 С. 55–56.
- ^ Бонда 2012, п. 171.
- ^ а б c Dennell & Roebroeks 2005, п. 1099.
- ^ а б Herries et al. 2020 г..
- ^ а б Кляйн 2005, п. 104.
- ^ а б Кляйн 2005, п. 93.
- ^ а б c Кляйн 2005, п. 101.
- ^ а б c Zhu et al. 2018 г., п. 608.
- ^ Кляйн 2005, п. 103.
- ^ Ларик и Чочон 1996, п. 1.
- ^ Чочон 2009, п. 910.
- ^ а б Кляйн 2005, п. 87.
- ^ а б c d Робертс 2018, п. 117.
- ^ а б c d е ж грамм час Кляйн 2005, п. 88.
- ^ Пагель и Бодмер 2004, п. 329.
- ^ Джайлз 2010, п. 326.
- ^ Давид-Барретт и Данбар, 2016 г..
- ^ Ракстон и Уилкинсон 2011, п. 20967.
- ^ Роджерс, Илтис и Вудинг, 2004 г. С. 105–108.
- ^ а б Кляйн 2005 С. 87–88.
- ^ а б Кляйн 2005, п. 89.
- ^ а б c Айелло и Уэллс 2002, п. 325.
- ^ а б Reno et al. 2003 г., п. 9404.
- ^ Kimbel & White 2017, п. 176.
- ^ а б Симпсон и др. 2008 г., п. 1090.
- ^ а б Coqueugniot et al. 2004 г. С. 299–302.
- ^ Каспари и Ли 2004.
- ^ а б c d е Айелло и Уэллс 2002, п. 326.
- ^ Ангар, Грин и Теафорд 2008 С. 208–228.
- ^ Бен-Дор и др. 2011 г..
- ^ Виллемс и Шайк 2017, п. 12.
- ^ а б Айелло и Уэллс 2002, п. 327.
- ^ Янссен, сентябрь и Гриффит, 2007 г., п. 308.
- ^ а б c Виллемс и Шайк 2017, п. 17.
- ^ Айелло и Данбар 1993, п. 184.
- ^ а б Айелло и Данбар 1993, п. 188.
- ^ а б Виллемс и Шайк 2017, п. 19.
- ^ Hatala et al. 2016 г., стр. 1–7.
- ^ а б Кляйн 2005, п. 94.
- ^ а б c Кляйн 2005, п. 95.
- ^ Новелл и Чанг 2009, п. 77.
- ^ Новелл и Чанг 2009, п. 83.
- ^ Новелл и Чанг 2009, п. 84.
- ^ Гоулетт 2016, п. 2.
- ^ Гоулетт 2016, п. 3.
- ^ а б Sandgathe & Berna 2017, п. S168.
- ^ а б Гоулетт 2016, п. 4.
- ^ Гоулетт 2016, п. 5.
- ^ Латимер и Оман 2001, п. A12.
- ^ Мейер, Векуа и Лордкипанидзе, 2006 г..
- ^ Schiess & Häusler, 2013 г., п. 365.
- ^ Schiess et al. 2014 г., п. 48.
Цитированная библиография
- Aiello, Leslie C .; Данбар, Р. И. М. (1993).«Размер неокортекса, размер группы и эволюция языка». Современная антропология. 34 (2): 184–193. Дои:10.1086/204160. S2CID 144347664.
- Aiello, Leslie C .; Уэллс, Джонатан К. К. (2002). «Энергетика и эволюция рода Гомо". Ежегодный обзор антропологии. 31: 323–338. Дои:10.1146 / annurev.anthro.31.040402.085403.
- Антон, Сьюзан С. (2003). "Естественная история человек прямоходящий". Ежегодник физической антропологии. 46: 126–170. Дои:10.1002 / ajpa.10399. PMID 14666536.
- Бааб, Карен Л. (2008). "Таксономические последствия изменения формы черепа у человек прямоходящий". Журнал эволюции человека. 54 (6): 827–847. Дои:10.1016 / j.jhevol.2007.11.003. PMID 18191986.
- Бонд, Нильс (2012). «Разнообразие гоминидов и мифы о« предках »». В Шильхабе, Тереза; Стьернфельт, Фредерик; Дьякон, Терренс (ред.). Эволюция символических видов. Springer. ISBN 978-94-007-2336-8.
- Бен-Дор, Мики; Гофер, Ави; Гершковиц, Израиль; Баркай, Ран (2011). "Человек-толстяк: Гибель человек прямоходящий и появление новой линии гомининов в среднем плейстоцене (около 400 тыс. лет) Левант ». PLOS ONE. 6 (12): e28689. Bibcode:2011PLoSO ... 628689B. Дои:10.1371 / journal.pone.0028689. ЧВК 3235142. PMID 22174868.
- Браун, Фрэнк; Харрис, Джон; Лики, Ричард; Уокер, Алан (1985). "Рано человек прямоходящий скелет из западного озера Туркана, Кения ". Природа. 316 (6031): 788–792. Bibcode:1985Натура.316..788Б. Дои:10.1038 / 316788a0. PMID 3929141. S2CID 4311887.
- Каспари, Рэйчел; Ли, Санг-Хи (2004). «Пожилой возраст становится обычным явлением в конце человеческой эволюции». PNAS. 101 (30): 10895–10900. Дои:10.1073 / pnas.0402857101. ЧВК 503716. PMID 15252198.
- Чочон, Рассел (2009). "Загадочная обезьяна плейстоценовой Азии" (PDF). Природа. 459 (7249): 910–911. Bibcode:2009Натура 459..910C. Дои:10.1038 / 459910a. PMID 19536242. S2CID 205047272.
- Coqueugniot, H .; Hublin, J.-J .; и другие. (2004). "Ранний рост мозга в человек прямоходящий и последствия для когнитивных способностей ". Природа. 431 (7006): 299–302. Bibcode:2004Натура 431..299C. Дои:10.1038 / природа02852. PMID 15372030. S2CID 4428043.
- Давид-Барретт, Тамаш; Данбар, Р. И. М. (2016). «Пересмотр двуногости и выпадения волос в эволюции человека: влияние высоты и расписания активности». Журнал эволюции человека. 94: 72–82. Дои:10.1016 / j.jhevol.2016.02.006. ЧВК 4874949. PMID 27178459.
- Деннелл, Робин; Робрукс, Уил (2005). «Азиатский взгляд на раннее расселение человека из Африки». Природа. 438 (7071): 1099–1104. Bibcode:2005Натура.438.1099D. Дои:10.1038 / природа04259. PMID 16371999. S2CID 4405913.
- Джайлз, Джеймс (2010). «Обнаженная любовь: эволюция безволосости человека». Биологическая теория. 5 (4): 326–336. Дои:10.1162 / BIOT_a_00062. S2CID 84164968.
- Гоулетт, Дж. А. Дж. (2016). «Открытие огня людьми: долгий и запутанный процесс». Философские труды B. 371 (20150164): 20150164. Дои:10.1098 / rstb.2015.0164. ЧВК 4874402. PMID 27216521.
- Hatala, Kevin G .; Роуч, Нил Т .; Ostrofsky, Kelly R .; Wunderlich, Roshna E .; Дингуолл, Хизер Л .; Вильмоар, Брайан А .; Грин, Дэвид Дж .; Харрис, Джон В. К .; Браун, Дэвид Р .; Ричмонд, Брайан Г. (2016). "Следы указывают на прямое свидетельство группового поведения и передвижения в человек прямоходящий". Научные отчеты. 6 (28766): 28766. Bibcode:2016НатСР ... 628766H. Дои:10.1038 / srep28766. ЧВК 4941528. PMID 27403790.
- Herries, Andy I. R .; и другие. (2020). "Современность Австралопитек, Парантроп, и рано человек прямоходящий в Южной Африке". Наука. 368 (6486): eaaw7293. Дои:10.1126 / science.aaw7293. HDL:11568/1040368. PMID 32241925. S2CID 214763272.
- Janssen, Marco A .; Сентябрь, Жанна М .; Гриффит, Кэмерон С. (2007). «Гоминиды, добывающие пищу в сложном ландшафте: могли Homo ergaster и Австралопитек бойцовский Удовлетворяют их потребности в калориях? »В Такахаши, Синго; Саллах, Давид; Рушье, Джульетта (ред.). Развитие социального моделирования: первый Всемирный конгресс. Springer. ISBN 978-4-431-73150-4.
- Kimbel, William H .; Белый, Тим Д. (2017). "Вариация, половой диморфизм и таксономия Австралопитек". В Grine, Фредерик Э. (ред.). Эволюционная история крепких австралопитеков. Рутледж. ISBN 978-0-202-36137-6.
- Кляйн, Ричард (2005). «Расселения гомининов в Старом Свете». В Крисе, Скарре (ред.). Человеческое прошлое: мировая предыстория и развитие человеческих обществ. Темза и Гудзон. ISBN 978-0500285312.
- Ларик, Рой; Чочон, Рассел (1996). "Возникновение в Африке и раннее азиатское расселение рода" Гомо". Американский ученый. 84 (6): 538–551. Bibcode:1996AmSci..84..538L.
- Латимер, Брюс; Оман, Джеймс С. (2001). «Осевая дисплазия в человек прямоходящий". Журнал эволюции человека. 40: A12.
- Meyer, Marc R .; Векуа, Абесалом; Лордкипанидзе, Давид (2006). "Язык и сочувствие в человек прямоходящий: поведение, предложенное современным спинным мозгом из Дманиси, но не Нариокотоме ". Цитировать журнал требует
| журнал =(помощь) - Новелл, апрель; Чанг, Мелани Ли (2009). «Дело против полового отбора как объяснение морфологии ручного топора» (PDF). Палеоантропология. 2009: 77–88.
- Пагель, Марк; Бодмер, Уолтер (2004). «Эволюция голого человека: культурные адаптации и гипотеза эктопаразитов». В Вассере, Соломон П. (ред.). Эволюционная теория и процессы: современные горизонты. Springer. Дои:10.1007/978-94-017-0443-4_17. ISBN 978-90-481-6457-8.
- Reno, Philip L .; Майндл, Ричард С .; Макколлум, Мелани А .; Лавджой, К. Оуэн (2003). "Половой диморфизм в Австралопитек афарский был подобен таковому у современных людей " (PDF). PNAS. 100 (16): 9404–9409. Bibcode:2003ПНАС..100.9404Р. Дои:10.1073 / pnas.1133180100. ЧВК 170931. PMID 12878734.
- Робертс, Алиса (2018). Эволюция: история человечества (Пересмотренная ред.). Dorling Kindersley Ltd. ISBN 978-0-2413-0431-0.
- Роджерс, Алан Р .; Илтис, Дэвид; Вудинг, Стивен (2004). «Генетические вариации в локусе MC1R и время, прошедшее с момента выпадения волос на теле человека». Современная антропология. 45 (1): 105–108. Дои:10.1086/381006.
- Ruxton, Graeme D .; Уилкинсон, Дэвид М. (2011). «Предотвращение перегрева и селекция как на выпадение волос, так и на двуногость у гомининов» (PDF). PNAS. 108 (52): 20965–20969. Bibcode:2011PNAS..10820965R. Дои:10.1073 / pnas.1113915108. ЧВК 3248486. PMID 22160694.
- Sandgathe, Dennis M .; Берна, Франческо (2017). "Огонь и род Гомо: Введение в Дополнение 16 ». Современная антропология. 58 (Приложение 16): S165 – S175. Дои:10.1086/691424. S2CID 164992270.
- Schiess, Regula; Хойслер, Мартин (2013). «Отсутствие скелетной дисплазии у мальчика из Нариокотома KNM-WT 15000 (человек прямоходящий) - переоценка врожденных патологий позвоночника ». Американский журнал физической антропологии. 150 (3): 365–374. Дои:10.1002 / ajpa.22211. PMID 23283736.
- Schiess, Regula; Бени, Томас; Рюли, Франк Дж .; Хойслер, Мартин (2014). «Возвращение к сколиозу в KNM-WT 15000» человек прямоходящий скелет " (PDF). Журнал эволюции человека. 67: 48–59. Дои:10.1016 / j.jhevol.2013.12.009. PMID 24491377.
- Шварц, Джеффри Х. (2000). «Таксономия Dmanisi Crania» (PDF). Наука. 289 (5476): 55–56. Дои:10.1126 / science.289.5476.55b. PMID 10928927. S2CID 23195350.
- Симпсон, Скотт У .; Куэйд, Джей; Левин, Наоми Э .; Батлер, Роберт; Дюпон-Ниве, Гийом; Эверетт, Мелани; Семав, Силеши (2008). "Женщина человек прямоходящий Таз из Гоны, Эфиопия ". Наука. 322 (5904): 1089–1092. Bibcode:2008Sci ... 322.1089S. CiteSeerX 10.1.1.710.7337. Дои:10.1126 / science.1163592. PMID 19008443. S2CID 22191315.
- Стрейт, Дэвид; Грин, Фредерик; Флигл, Джон (2015). "Анализ филогении гомининов: кладистический подход" (PDF). В Хенке, Винфрид; Таттерсолл, Ян (ред.). Справочник по палеоантропологии (2-е изд.). Springer. Дои:10.1007/978-3-642-39979-4_58. ISBN 978-3-642-39979-4.
- Таттерсолл, Ян (2013). "Homo ergaster и его современники » (PDF). В Хенке, Винфрид; Таттерсолл, Ян (ред.). Справочник по палеоантропологии. Springer. ISBN 978-3-642-39978-7.
- Ungar, Peter S .; Грин, Фредерик Э .; Тефорд, Марк Ф. (2008). «Диета в начале Гомо: Обзор доказательств и новая модель адаптивной универсальности ». Ежегодный обзор антропологии. 35 (1): 208–228.
- Виллемс, Эрик П .; ван Шайк, Карел П. (2017). "Социальная организация Homo ergaster: Выводы из реакции против хищников у современных приматов ". Журнал эволюции человека. 109: 11–21. Дои:10.1016 / j.jhevol.2017.05.003. PMID 28688456.
- Ван, Вэйцзе; Кромптон, Робин Н .; Кэри, Таня С .; Гюнтер, Майкл М .; Ли, Ю; Сэвидж, Рассел; Продавцы, Уильямс I. (2004). «Сравнение скелетно-мышечных моделей с обратной динамикой AL 288-1. Австралопитек афарский и KNM-WT 15000 Homo ergaster современному человеку, что имеет значение для эволюции двуногости ". Журнал эволюции человека. 47 (6): 453–478. Дои:10.1016 / j.jhevol.2004.08.007. PMID 15566947.
- Вуд, Бернард; Коллард, Марк (1999). "Человеческий род". Наука. 284 (5411): 65–71. Bibcode:1999Научный ... 284 ... 65.. Дои:10.1126 / наука.284.5411.65. PMID 10102822.
- Чжу, Чжаоюй; Деннелл, Робин; Хуанг, Вэйвэнь; Ву, Йи; Цю, Шифань; Ян, Шися; Рао, Чжиго; Хоу, Ямэй; Се, Цзюбинь; Хан, Цзянвэй; Оуян, Тинпин (2018). «Гомининское заселение китайского Лессового плато примерно 2,1 миллиона лет назад». Природа. 559 (7715): 608–612. Bibcode:2018Натура.559..608Z. Дои:10.1038 / s41586-018-0299-4. PMID 29995848. S2CID 49670311.
внешняя ссылка
- Homo ergaster; Австралийский музей
- Homo ergaster; Издательство Милна - История нашего племени: Гоминини
- Homo ergaster; Происхождение - Изучение летописи окаменелостей - Фонд Брэдшоу
- Homo ergaster; Ресурсы по eFossils
- Генеалогическое древо происхождения человека; Центр обучения ДНК
| Таксономия (Гоминины ) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Предки |
| ||||||||||||||||||||||||||
| Модели |
| ||||||||||||||||||||||||||
| Сроки | |||||||||||||||||||||||||||
| Другие |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||