Эволюционная генетика человека - Human evolutionary genetics
Эволюционная генетика человека изучает, как один человеческий геном отличается от другого генома человека, эволюционным прошлым, которое привело к возникновению генома человека, и его текущими последствиями. Различия между геномами имеют антропологический, медицинский, исторические и судебно-медицинский последствия и приложения. Генетические данные могут дать важную информацию о эволюция человека.
Происхождение обезьян
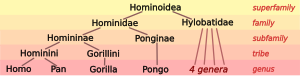
Биологи классифицируют люди, наряду с некоторыми другими разновидность, в качестве большие обезьяны (виды в семья Гоминиды ). Живущие гоминиды включают два разных вида: шимпанзе (в бонобо, Пан панискус, а обыкновенный шимпанзе, Пан троглодиты), два вида горилла (в западная горилла, Горилла горилла, а восточная горилла, Горилла грауэри), и два вида орангутанг (в Борнейский орангутанг, Pongo pygmaeus, а Суматранский орангутанг, Pongo abelii). Великие обезьяны с семьей Hylobatidae гиббонов образуют надсемейство Гоминоидея обезьян.
Обезьяны, в свою очередь, относятся к примат порядка (> 400 видов), а также Обезьяны Старого Света, то Обезьяны Нового Света, и другие. Данные из обоих митохондриальная ДНК (мтДНК) и ядерная ДНК (яДНК) указывают на то, что приматы принадлежат к группе Euarchontoglires, вместе с Rodentia, Зайцеобразные, Dermoptera, и Scandentia.[1] Это дополнительно поддерживается Alu-подобным короткие вкрапленные ядерные элементы (SINE), которые были обнаружены только у членов Euarchontoglires.[2]
Филогенетика
А филогенетическое дерево обычно происходит от ДНК или же белок последовательности от населения. Часто, митохондриальная ДНК или же Y-хромосома последовательности используются для изучения демографии древнего человека. Эти одно-локус источники ДНК не рекомбинировать и почти всегда наследуются от одного родителя, за исключением только одного известного исключения в мтДНК.[3] Люди из более близких географических регионов обычно более похожи, чем люди из более отдаленных регионов. Расстояние на филогенетическом дереве можно приблизительно использовать для обозначения:
- Генетическая дистанция. Генетическая разница между людьми и шимпанзе составляет менее 2%,[4] или в три раза больше, чем различия среди современных людей (оценка 0,6%).[5]
- Временная удаленность последнего общего предка. В митохондриальный самый последний общий предок современных людей, по оценкам, жили примерно 160000 лет назад,[6] последние общие предки людей и шимпанзе примерно 5-6 миллионов лет назад.[7]
Видообразование человека и африканских обезьян
Отделение людей от их ближайших родственников, нечеловеческих обезьян (шимпанзе и горилл), широко изучается более века. Были рассмотрены пять основных вопросов:
- Какие обезьяны являются нашими ближайшими предками?
- Когда произошли разлуки?
- Что это было эффективная численность населения общего предка до раскола?
- Существуют ли следы популяционной структуры (субпопуляции), предшествующие видообразованию, или частичная примесь, следующая за ним?
- Каковы были конкретные события (включая слияние хромосом 2a и 2b) до и после разделения?
Общие наблюдения
Как обсуждалось ранее, разные части генома демонстрируют различное расхождение последовательностей между разными гоминоиды. Также было показано, что расхождение последовательностей ДНК человека и шимпанзе сильно различается. Например, расхождение последовательности варьируется от 0% до 2,66% между некодирующими и неповторяющимися геномный регионы людей и шимпанзе.[8] Процент нуклеотидов в геноме человека (hg38), имевших однозначное соответствие в геноме шимпанзе (pantro6), составил 84,38%. Кроме того, деревья генов, полученные путем сравнительного анализа сегментов ДНК, не всегда соответствуют дереву видов. Подводя итоги:
- Расхождение последовательностей значительно различается у людей, шимпанзе и горилл.
- По большинству последовательностей ДНК люди и шимпанзе кажутся наиболее близкими родственниками, но некоторые указывают на человека-гориллу или шимпанзе-гориллу. клады.
- Был секвенирован геном человека и геном шимпанзе. Люди имеют 23 пары хромосом, а шимпанзе, гориллы и орангутаны есть 24. Хромосома человека 2 представляет собой слияние двух хромосом 2a и 2b, которые остались отдельными у других приматов.[9]
Время расхождения
Большой интерес представляет время расхождения человека с другими обезьянами. В одном из первых молекулярных исследований, опубликованных в 1967 году, измерялись иммунологические расстояния (ID) между разными приматами.[10] В основном в исследовании измерялась сила иммунологического ответа, который антиген от одного вида (человеческий альбумин) индуцирует в иммунной системе другого вида (человека, шимпанзе, гориллы и Обезьяны Старого Света ). Близкородственные виды должны иметь аналогичные антигены и, следовательно, более слабый иммунологический ответ на антигены друг друга. Иммунологический ответ вида на его собственные антигены (например, от человека к человеку) был установлен равным 1.
ID между людьми и гориллами был определен равным 1,09, а между людьми и шимпанзе - 1,14. Однако расстояние до шести разных обезьян Старого Света было в среднем 2,46, что указывает на то, что африканские обезьяны более тесно связаны с людьми, чем с обезьянами. Авторы полагают, что время расхождения между обезьянами Старого Света и гоминоидами составляет 30 миллионов лет назад (MYA), на основании данных по ископаемым, и считается, что иммунологическое расстояние растет с постоянной скоростью. Они пришли к выводу, что время расхождения людей и африканских обезьян составляет примерно ~ 5 млн лет назад. Это был удивительный результат. Большинство ученых того времени считали, что люди и человекообразные обезьяны разошлись намного раньше (> 15 млн лет назад).
Горилла с точки зрения идентификации была ближе к человеку, чем к шимпанзе; однако разница была настолько незначительной, что трихотомия не может быть решен с уверенностью. Более поздние исследования, основанные на молекулярной генетике, позволили разрешить трихотомию: шимпанзе - это филогенетически ближе к людям, чем к гориллам. Однако некоторые времена расхождения, оцененные позже (с использованием гораздо более сложных методов молекулярной генетики), существенно не отличаются от самой первой оценки 1967 года, но недавняя статья[11] ставит это в 11–14 MYA.
Время расхождения и эффективный размер популяции предков

Современные методы определения времени расхождения используют выравнивание последовательностей ДНК и молекулярные часы. Обычно молекулярные часы калибруют, исходя из предположения, что орангутанг отделился от африканских обезьян (включая людей) за 12–16 лет. Некоторые исследования также включают некоторых обезьян старого мира и устанавливают время их расхождения от гоминоидов на уровне 25-30 млн лет. Обе точки калибровки основаны на очень небольшом количестве данных об ископаемых и подвергались критике.[12]
Если эти даты будут пересмотрены, время расхождения, оцененное на основе молекулярных данных, также изменится. Однако маловероятно, что относительное время расхождения изменится. Даже если мы не можем точно определить время абсолютного расхождения, мы можем быть уверены, что время расхождения между шимпанзе и человеком примерно в шесть раз короче, чем между шимпанзе (или людьми) и обезьянами.
Одно исследование (Такахата и другие., 1995) использовали 15 последовательностей ДНК из разных областей генома человека и шимпанзе и 7 последовательностей ДНК человека, шимпанзе и гориллы.[13] Они определили, что шимпанзе более тесно связаны с людьми, чем гориллы. Используя различные статистические методы, они оценили время расхождения между человеком и шимпанзе в 4,7 млн лет, а время расхождения между горилл и человеком (и шимпанзе) - 7,2 млн лет.
Кроме того, они оценили эффективная численность населения общего предка людей и шимпанзе ~ 100000. Это было несколько удивительно, поскольку в настоящее время эффективная численность населения оценивается всего в ~ 10 000 человек. Если это правда, это означает, что человеческое происхождение испытало бы огромное уменьшение своего эффективного размера популяции (и, следовательно, генетического разнообразия) в своей эволюции. (видеть Теория катастрофы Тоба )

Другое исследование (Chen & Li, 2001) секвенировало 53 неповторяющихся межгенных сегмента ДНК из человек, шимпанзе, горилла и орангутанг.[8] Когда последовательности ДНК были объединены в одну длинную последовательность, сгенерированный присоединение к соседу дерево поддержало Гомо-Сковорода clade со 100% бутстрапом (то есть люди и шимпанзе являются ближайшими родственными видами из четырех). Когда три вида довольно тесно связаны друг с другом (например, человек, шимпанзе и горилла), деревья, полученные из данных последовательности ДНК, могут не совпадать с деревом, представляющим видообразование (дерево видов).
Более короткий межузловой промежуток времени (TВ) более распространены неконгруэнтные генные деревья. Эффективная численность популяции (Nе) интернодальной популяции определяет, как долго генетические линии сохраняются в популяции. Чем выше эффективный размер популяции, тем больше несовместимых генов. Следовательно, если известен межузловой промежуток времени, можно рассчитать эффективный размер популяции предков общего предка людей и шимпанзе.
Когда каждый сегмент анализировался индивидуально, 31 подтвердил Гомо-Сковорода clade, 10 поддержали Гомо-Горилла clade, а 12 поддержали Сковорода-Горилла клады. Используя молекулярные часы, авторы подсчитали, что гориллы разделились в первые 6,2–8,4 млн лет назад, а шимпанзе и люди разделились на 1,6–2,2 миллиона лет спустя (межузловой промежуток времени) 4,6–6,2 млн лет назад. Промежуток времени между узлами полезен для оценки эффективного размера популяции предков общего предка людей и шимпанзе.
А скупой анализ показал, что 24 локуса поддерживали Гомо-Сковорода clade, 7 поддержали Гомо-Горилла clade, 2 поддержали Сковорода-Горилла clade и 20 не дали разрешения. Дополнительно они взяли из баз данных 35 локусов, кодирующих белок. Из них 12 поддержали Гомо-Сковорода клады, 3 Гомо-Горилла клады, 4 Сковорода-Горилла clade и 16 не дали разрешения. Следовательно, только ~ 70% из 52 локусов, которые дали разрешение (33 межгенных, 19 кодирующих белков), поддерживают «правильное» дерево видов. Исходя из доли локусов, не поддерживающих дерево видов, и интернодального промежутка времени, который они оценили ранее, эффективная популяция общего предка человека и шимпанзе оценивается в ~ 52 000–96 000. Это значение не так высоко, как это из первого исследования (Такахата), но все же намного выше, чем нынешняя эффективная численность населения.
В третьем исследовании (Янг, 2002) использовался тот же набор данных, что и Чен и Ли, но по оценке предков эффективное население «всего» от ~ 12 000 до 21 000, используя другой статистический метод.[14]
Генетические различия между людьми и другими человекообразными обезьянами
Выравниваемые последовательности в геномах человека и шимпанзе отличаются примерно на 35 миллионов однонуклеотидных замен. Кроме того, около 3% полных геномов различаются делециями, вставками и дупликациями.[15]
Поскольку частота мутаций относительно постоянна, примерно половина этих изменений произошла в человеческой линии. Лишь очень крошечная часть этих фиксированных различий дала начало различным фенотипам человека и шимпанзе, и их обнаружение является большой проблемой. Подавляющее большинство различий нейтральны и не влияют на фенотип.[нужна цитата ]
Молекулярная эволюция может действовать по-разному: через эволюцию белка, потерю генов, дифференциальную регуляцию генов и эволюцию РНК. Считается, что все они сыграли определенную роль в эволюции человека.
Потеря гена
Многие различные мутации могут инактивировать ген, но немногие изменят его функцию определенным образом. Таким образом, инактивационные мутации будут легко доступны для отбора. Таким образом, потеря генов может быть обычным механизмом эволюционной адаптации (гипотеза «меньше значит больше»).[16]
80 генов были потеряны в человеческой линии после отделения от последнего общего предка с шимпанзе. 36 из них были для обонятельные рецепторы. Гены, участвующие в хеморецепции и иммунном ответе, чрезмерно представлены.[17] Другое исследование показало, что 86 генов были потеряны.[18]
Ген кератина волос KRTHAP1
Ген волос типа I кератин был потерян в человеческом роде. Кератины - основной компонент волос. У людей все еще есть девять функциональных генов кератина волос I типа, но потеря этого гена могла вызвать истончение волос на теле человека. Основываясь на предположении о постоянных молекулярных часах, исследование предсказывает, что потеря гена произошла относительно недавно в эволюции человека - менее 240000 лет назад, но и последовательность Виндия неандертальца, и денисовская последовательность с высоким охватом содержат те же преждевременные стоп-кодоны, что и современные человека и, следовательно, датировка должна быть более 750 000 лет назад. [19]
Ген миозина MYH16
Стедман и другие. (2004) заявили, что потеря саркомера миозин ген MYH16 в человеческом роде привело к меньшим жевательные мышцы. Они подсчитали, что мутация, которая привела к инактивации (делеция двух пар оснований), произошла 2,4 миллиона лет назад, до появления Homo ergaster / erectus в Африке. Последующий период ознаменовался сильным увеличением черепная емкость, способствуя предположению, что потеря гена могла устранить эволюционное ограничение на размер мозга у представителей рода Гомо.[20]
По другой оценке, потеря гена MYH16 произошла 5,3 миллиона лет назад, задолго до этого. Гомо появившийся.[21]
Другой
- CASPASE12, цистеинил аспартат протеиназа. Предполагается, что потеря этого гена снизила летальность бактериальной инфекции у людей.[17]
Добавление гена
Сегментарные дупликации (SD или LCR ) сыграли роль в создании новых генов приматов и формировании генетической изменчивости человека.
Вставки ДНК человека
Когда человек геном сравнивали с геномами пяти сравниваемых видов приматов, включая шимпанзе, горилла, орангутанг, gibbon и macaque, было обнаружено, что существует около 20000 специфичных для человека вставок, которые считаются регулирующими. Хотя большинство вставок кажутся нейтральными к приспособленности, небольшое количество было идентифицировано в положительно выбранных генах, показывающих ассоциации с нейронными фенотипами, а некоторые - с фенотипами, связанными с зубами и сенсорным восприятием. Эти находки намекают на кажущуюся важную роль специфичных для человека вставок в недавней эволюции человека.[22]
Выбор давления
Человеческие ускоренные области - это области генома, которые различаются между людьми и шимпанзе в большей степени, чем можно объяснить генетическим дрейфом с течением времени, поскольку у этих двух видов был общий предок. Эти регионы демонстрируют признаки естественного отбора, ведущего к эволюции отчетливо человеческих черт. Два примера: HAR1F, который, как полагают, связан с развитием мозга и HAR2 (a.k.a. HACNS1 ), которые могли сыграть роль в развитии противопоставленный большой палец.
Также была выдвинута гипотеза, что большая часть различий между людьми и шимпанзе связана с регуляция экспрессии генов а не различия в самих генах. Анализ консервативные некодирующие последовательности, которые часто содержат функциональные и, таким образом, положительно выбранные регуляторные области, обращаются к этой возможности.[23]
Расхождение последовательностей между людьми и обезьянами
Когда черновая последовательность обыкновенного шимпанзе (Пан троглодиты) геном был опубликован летом 2005 года, 2400 миллионов оснований (из ~ 3160 миллионов оснований) были секвенированы и собраны достаточно хорошо, чтобы их можно было сравнить с геномом человека.[15] 1,23% из этого секвенирования отличались заменами на одно основание. Считалось, что 1,06% или менее из них представляют фиксированные различия между видами, а остальные являются вариантами участков у людей или шимпанзе. Другой тип различия, называемый инделы (вставки / удаления) составляли гораздо меньше различий (на 15% больше), но вносили ~ 1,5% уникальной последовательности в каждый геном, поскольку каждая вставка или удаление может включать от одного основания до миллионов оснований.[15]
Сопутствующий документ изучен сегментарные дупликации в двух геномах,[24] чьи вставки и удаления в геном составляют большую часть последовательности indel. Они обнаружили, что всего 2,7% эухроматической последовательности были дифференциально дублированы в той или иной линии.
| Locus | Человек-шимпанзе | Человек-горилла | Человек-Орангутанг |
|---|---|---|---|
| Элементы Alu | 2 | - | - |
| Некодирование (Chr. Y) | 1.68 ± 0.19 | 2.33 ± 0.2 | 5.63 ± 0.35 |
| Псевдогены (аутосомные) | 1.64 ± 0.10 | 1.87 ± 0.11 | - |
| Псевдогены (Chr. X) | 1.47 ± 0.17 | - | - |
| Некодирование (аутосомно) | 1.24 ± 0.07 | 1.62 ± 0.08 | 3.08 ± 0.11 |
| Гены (Ks) | 1.11 | 1.48 | 2.98 |
| Интроны | 0.93 ± 0.08 | 1.23 ± 0.09 | - |
| Xq13.3 | 0.92 ± 0.10 | 1.42 ± 0.12 | 3.00 ± 0.18 |
| Итого по Х-хромосоме | 1.16 ± 0.07 | 1.47 ± 0.08 | - |
| Гены (Kа) | 0.8 | 0.93 | 1.96 |
Расхождение последовательностей обычно имеет следующий характер: Человек-Шимпанзе <Человек-Горилла << Человек-Орангутанг, что подчеркивает близкое родство между людьми и африканскими обезьянами. Элементы Alu быстро расходятся из-за высокой частоты CpG динуклеотиды, которые мутируют примерно в 10 раз чаще, чем средний нуклеотид в геноме. Скорость мутации выше у мужчин линия зародыша, поэтому расхождение в Y-хромосома - который наследуется исключительно от отца - выше, чем в аутосомы. В Х хромосома наследуется через женскую зародышевую линию в два раза чаще, чем через мужскую зародышевую линию, поэтому расхождение последовательностей несколько ниже. Дивергенция последовательности области Xq13.3 удивительно низка между людьми и шимпанзе.[25]
Мутации, изменяющие аминокислотную последовательность белков (Kа) являются наименее распространенными. Фактически ~ 29% всех ортологичных белков человека и шимпанзе идентичны. Типичный белок отличается всего двумя аминокислотами.[15]Показатели расхождения последовательностей, показанные в таблице, принимают только различия замещения, например, от A (аденин ) в G (гуанин ), в учетную запись. Однако последовательности ДНК могут также отличаться вставками и делециями (инделы ) баз. Обычно они удаляются из выравниваний перед вычислением дивергенции последовательностей.
Генетические различия между современными людьми и неандертальцами
Международная группа ученых завершила набросок последовательности Неандерталец генома в мае 2010 года. Результаты указывают на некоторые размножение между современными людьми (Homo sapiens) и неандертальцы (Homo neanderthalensis), поскольку геномы людей неафриканского происхождения имеют на 1–4% больше общего с неандертальцами, чем геномы африканцев к югу от Сахары. Неандертальцы и большинство современных людей разделяют непереносимость лактозы вариант лактаза ген, кодирующий фермент, который не может расщеплять лактозу в молоке после отъема. Современные люди и неандертальцы также разделяют FOXP2 вариант гена, связанный с развитием мозга и речью у современных людей, что указывает на то, что неандертальцы могли говорить. Шимпанзе имеют два аминокислотных различия в FOXP2 по сравнению с человеческим и неандертальским FOXP2.[26][27][28]
Генетические различия между современными людьми
Х. сапиенс считается, что он появился около 300 000 лет назад. Он разошелся по всей Африке, а после 70000 лет назад по всей Евразии и Океании. Исследование 2009 года выявило 14 «групп предкового населения», наиболее удаленными из которых являются Народ сан Южной Африки.[29][30]
Благодаря их быстрому распространению в различных климатических зонах, и особенно с появлением новых источников пищи с приручение крупного рогатого скота и развитие сельского хозяйства, человеческое население подверглось значительному селективное давление с момента их разгона. Например, Выходцы из Восточной Азии были обнаружены отделенными от Европиды рядом концентрированных аллелей, указывающих на давление отбора, включая варианты ЕДАР, ADH1B, ABCC1, и ALDH2genes Восточноазиатские типы ADH1B, в частности, связаны с приручение риса и, таким образом, возникла бы после развития рисоводства примерно 10 000 лет назад.[31] Некоторые фенотипические черты, характерные для жителей Восточной Азии, связаны с единственной мутацией ЕДАР ген, датированный c. 35000 лет назад.[32]
По состоянию на 2017 год[Обновить], База данных одиночного нуклеотидного полиморфизма (dbSNP ), в котором перечислены SNP и другие варианты, перечислено в общей сложности 324 миллиона вариантов, обнаруженных в секвенированных геномах человека.[33]Нуклеотидное разнообразие, средняя доля нуклеотидов, которые различаются между двумя людьми, оценивается в пределах от 0,1% до 0,4% для современных людей (по сравнению с 2% между людьми и шимпанзе).[34][35]Это соответствует различиям в геномах нескольких миллионов сайтов; то Проект 1000 геномов аналогичным образом было обнаружено, что «типичный [индивидуальный] геном отличается от эталонного генома человека на 4,1-5,0 миллионов сайтов… затрагивающих 20 миллионов оснований последовательности».[36]
В феврале 2019 года ученые обнаружили доказательства, основанные на генетические исследования с помощью искусственный интеллект (AI), которые предполагают существование неизвестного вида предков человека, а не Неандерталец, Денисовский или гибрид человека (например, Денни (гибрид гоминин) ), в геном из современные люди.[37][38]
Научные исследования
В марте 2019 года китайские ученые сообщили об установке человеческого мозга. Ген MCPH1 в лабораторию макаки резус, в результате чего трансгенные обезьяны лучше работают и быстрее отвечают на «тесты краткосрочной памяти, включающие соответствие цвета и формы», по сравнению с контрольными нетрансгенными обезьянами, по мнению исследователей.[39][40]
Смотрите также
- Археогенетика Ближнего Востока
- Проект генома шимпанзе
- FOXP2 и эволюция человека
- Генетика и археогенетика Южной Азии
- Генетическая история Европы
- Генетическая история коренных народов Америки
- Генетическая история Италии
- Генетическая история Британских островов
- Гоминины
- Список гаплогрупп исторических и известных личностей
- Путешествие человека: генетическая одиссея
- Гаплогруппы Y-ДНК по этническим группам
- Раса и генетика
- Недавняя эволюция человека
Рекомендации
- ^ Мерфи, W.J .; Eizirik, E .; О'Брайен, С.Дж .; Madsen, O .; Скалли, М .; Douady, C.J .; Teeling, E .; Ryder, O.A .; Стэнхоуп, M.J .; de Jong, W.W .; Спрингер, М. (2001). «Разрешение раннего излучения плаценты млекопитающих с использованием байесовской филогенетики». Наука. 294 (5550): 2348–2351. Bibcode:2001Научный ... 294.2348M. Дои:10.1126 / science.1067179. PMID 11743200. S2CID 34367609.
- ^ Kriegs, J.O .; Чураков, Г .; Kiefmann, M .; Jordan, U .; Brosius, J .; Шмитц, Дж. (2006). «Ретропозиционные элементы как архив эволюционной истории плацентарных млекопитающих». ПЛОС Биол. 4 (4): e91. Дои:10.1371 / journal.pbio.0040091. ЧВК 1395351. PMID 16515367.
- ^ Шварц М., Виссинг Дж. (2002). «Отцовское наследование митохондриальной ДНК». N Engl J Med. 347 (8): 576–580. Дои:10.1056 / NEJMoa020350. PMID 12192017.
- ^ "Хромосома 2 человека" PBS.
- ^ По состоянию на 2015 год типичное различие между геномами двух индивидуумов оценивалось в 20 миллионов пар оснований (или 0,6% от общего количества 3,2 миллиарда пар оснований): «Типичный [индивидуальный] геном отличается от эталонного генома человека на 4,1 миллиона. до 5,0 миллионов сайтов, [...] затрагивающих 20 миллионов оснований последовательности »Аутон А., Брукс Л.Д., Дурбин Р.М., Гаррисон Е.П., Кан Х.М., Корбел Дж.О. и др. (Октябрь 2015 г.). «Глобальный справочник генетических вариаций человека». Природа. 526 (7571): 68–74. Bibcode:2015 Натур.526 ... 68 т. Дои:10.1038 / природа15393. ЧВК 4750478. PMID 26432245.
- ^ «134 - 188 тыс. Лет назад»: Fu Q, Mittnik A, Johnson PL, Bos K, Lari M, Bollongino R, Sun C, Giemsch L, Schmitz R, Burger J, Ronchitelli AM, Martini F, Cremonesi RG, Svoboda J, Bauer P, Caramelli D, Castellano S , Reich D, Pääbo S, Krause J (21 марта 2013 г.). «Пересмотренная шкала времени эволюции человека на основе древних митохондриальных геномов». Текущая биология. 23 (7): 553–59. Дои:10.1016 / j.cub.2013.02.044. ЧВК 5036973. PMID 23523248..
- ^ Паттерсон Н., Рихтер Д. Д., Гнерре С., Ландер Э. С., Рейх Д. (2006). «Генетические свидетельства сложного видообразования человека и шимпанзе». Природа. 441 (7097): 1103–8. Bibcode:2006 Натур.441.1103П. Дои:10.1038 / природа04789. PMID 16710306. S2CID 2325560.
- ^ а б c Chen, F.C .; Ли, W.H. (2001). «Геномные расхождения между людьми и другими гоминоидами и эффективный размер популяции общего предка людей и шимпанзе». Am J Hum Genet. 68 (2): 444–456. Дои:10.1086/318206. ЧВК 1235277. PMID 11170892.
- ^ Кен Миллер в Китцмиллер против Дувра протоколы судебных заседаний.
- ^ Сарич, В.М .; Уилсон, A.C. (1967). «Иммунологическая шкала времени для эволюции гоминидов». Наука. 158 (3805): 1200–1203. Bibcode:1967Sci ... 158.1200S. Дои:10.1126 / science.158.3805.1200. PMID 4964406. S2CID 7349579.
- ^ Венн, Оливер; Тернер, Исаак; Мэтисон, Иэн; де Гроот, Натасья; Бонтроп, Рональд; Маквин, Гил (июнь 2014 г.). «Сильное мужское предубеждение приводит к мутации зародышевой линии у шимпанзе». Наука. 344 (6189): 1272–1275. Bibcode:2014Научный ... 344.1272V. Дои:10.1126 / science.344.6189.1272. ЧВК 4746749. PMID 24926018.
- ^ Yoder, A.D .; Янг, З. (1 июля 2000 г.). «Оценка дат видообразования приматов с использованием локальных молекулярных часов». Мол Биол Эвол. 17 (7): 1081–1090. Дои:10.1093 / oxfordjournals.molbev.a026389. PMID 10889221.
- ^ Takahata, N .; Satta, Y .; Кляйн, Дж. (1995). «Время расхождения и размер популяции в родословной, ведущей к современным людям». Theor Popul Biol.. 48 (2): 198–221. Дои:10.1006 / tpbi.1995.1026. PMID 7482371.
- ^ Янг, З. (1 декабря 2002 г.). «Вероятность и Байесовская оценка размеров предковой популяции гоминоидов с использованием данных из нескольких локусов» (аннотация страница). Генетика. 162 (4): 1811–1823. ЧВК 1462394. PMID 12524351.
- ^ а б c d Секвенирование шимпанзе; Консорциум анализа (2005 г.). «Исходная последовательность генома шимпанзе и сравнение с геномом человека». Природа. 437 (7055): 69–87. Bibcode:2005Натура 437 ... 69.. Дои:10.1038 / природа04072. PMID 16136131.
- ^ Олсон, М. (1999). «Когда меньше - значит больше: потеря генов как двигатель эволюционных изменений». Am J Hum Genet. 64 (1): 18–23. Дои:10.1086/302219. ЧВК 1377697. PMID 9915938.
- ^ а б Ван, X .; Grus, W.E .; Чжан, Дж. (2006). «Потери генов при человеческом происхождении». ПЛОС Биол. 4 (3): e52. Дои:10.1371 / journal.pbio.0040052. ЧВК 1361800. PMID 16464126.
- ^ Демут, Джеффри П .; Би, Тиджл Де; Stajich, Jason E .; Кристианини, Нелло; Хан, Мэтью В. (декабрь 2006 г.). Боревиц, Джастин (ред.). "Эволюция генных семейств млекопитающих". PLOS ONE. 1 (1): e85. Bibcode:2006PLoSO ... 1 ... 85D. Дои:10.1371 / journal.pone.0000085. ЧВК 1762380. PMID 17183716.
- ^ Winter, H .; Langbein, L .; Krawczak, M .; Cooper, D.N .; Suarez, L.F.J .; Rogers, M.A .; Praetzel, S .; Heidt, P.J .; Швейцер, Дж. (2001). «Псевдоген кератина волос человека типа I phihHaA имеет функциональные ортологи у шимпанзе и горилл: свидетельство недавней инактивации гена человека после дивергенции Pan-Homo». Hum Genet. 108 (1): 37–42. Дои:10.1007 / s004390000439. PMID 11214905. S2CID 21545865.
- ^ Stedman, H.H .; Kozyak, B.W .; Нельсон, А .; Thesier, D.M .; Su, L.T .; Low, D.W .; Bridges, C.R .; Shrager, J.B .; Purvis, N.M .; Митчелл, М.А. (2004). «Мутация гена миозина коррелирует с анатомическими изменениями человеческого происхождения». Природа. 428 (6981): 415–418. Bibcode:2004Натура.428..415S. Дои:10.1038 / природа02358. PMID 15042088. S2CID 4304466.
- ^ Perry, G.H .; Verrelli, B.C .; Стоун, A.C. (2005). «Сравнительный анализ раскрывает сложную историю молекулярной эволюции MYH16 человека». Мол Биол Эвол. 22 (3): 379–382. Дои:10.1093 / molbev / msi004. PMID 15470226.
- ^ Hellen, Elizabeth H.B .; Керн, Эндрю Д. (2015-04-01). «Роль вставок ДНК в фенотипической дифференциации между людьми и другими приматами». Геномная биология и эволюция. 7 (4): 1168–1178. Дои:10.1093 / gbe / evv012. ISSN 1759-6653. ЧВК 4419785. PMID 25635043.
- ^ Bird, Christine P .; Лю, Морин; и другие. (2007). «Быстро развивающиеся некодирующие последовательности в геноме человека». Геномная биология. 8 (6): R118. Дои:10.1186 / gb-2007-8-6-r118. ЧВК 2394770. PMID 17578567.
- ^ Cheng, Z .; Ventura, M .; Она, X .; Хаитович, П .; Могилы, Т .; Osoegawa, K .; Церковь, D; Pieter DeJong, P .; Wilson, R.K .; Paabo, S .; Рокки, М; Эйхлер, Э. Э. (2005). «Полногеномное сравнение недавних сегментарных дупликаций шимпанзе и человека». Природа. 437 (1 сентября 2005 г.): 88–93. Bibcode:2005Натура 437 ... 88С. Дои:10.1038 / природа04000. PMID 16136132. S2CID 4420359.
- ^ Kaessmann, H .; Heissig, F .; von Haeseler, A .; Паабо, С. (1999). «Вариация последовательности ДНК в некодирующей области с низкой рекомбинацией на Х-хромосоме человека». Нат Жене. 22 (1): 78–81. Дои:10.1038/8785. PMID 10319866. S2CID 9153915.
- ^ Сэй, Тина Хесман (2009). «История первая: команда расшифровывает ДНК неандертальцев: черновик генома может раскрыть секреты эволюции человека». Новости науки. 175 (6): 5–7. Дои:10.1002 / scin.2009.5591750604.
- ^ Грин, Ричард Э .; Краузе; Бриггс; Маричич; Стензель; Кирхер; Паттерсон; Ли; Чжай; Фриц; Хансен; Дюран; Маласпинас; Дженсен; Маркес-Боне; Алкан; Прюфер; Мейер; Бурбано; Хороший; Шульц; Аксиму-Петри; Butthof; Хёбер; Хёффнер; Зигемунд; Вейманн; Нусбаум; Посадочный модуль; и другие. (07.05.2010). «Черновик последовательности генома неандертальца». Наука. 328 (5979): 710–722. Bibcode:2010Sci ... 328..710G. Дои:10.1126 / science.1188021. ЧВК 5100745. PMID 20448178.
- ^ "НЕАНДЕРТАЛЫ ЖИВЫ!". блог Джона Хоукса. Получено 2010-12-31.
- ^ Тишкофф, С.А.; Рид, FA .; Friedlaender, FR .; Ehret, C .; Ranciaro, A .; Froment, A .; Hirbo, JB .; Awomoyi, AA .; и другие. (Май 2009 г.). «Генетическая структура и история африканцев и афроамериканцев». Наука. 324 (5930): 1035–44. Bibcode:2009Sci ... 324.1035T. Дои:10.1126 / science.1172257. ЧВК 2947357. PMID 19407144.
- ^ BBC World News: "Генетические секреты Африки раскрыты", 1 мая 2009 г.
- ^ Peng, Y .; и другие. (2010). «Полиморфизм ADH1B Arg47His в популяциях Восточной Азии и расширение одомашнивания риса в истории». BMC Эволюционная биология. 10: 15. Дои:10.1186/1471-2148-10-15. ЧВК 2823730. PMID 20089146.
- ^ Мутация затрагивает потовые железы, зубы, толщину волос и ткань груди. Камберов; и другие. (2013). «Моделирование недавней эволюции человека на мышах путем выражения выбранного варианта EDAR». Клетка. 152 (4): 691–702. Дои:10.1016 / j.cell.2013.01.016. ЧВК 3575602. PMID 23415220. Восточноазиатские физические особенности связаны с 35000-летней мутацией, NYT, 14 февраля 2013 г.
- ^ NCBI (2017-05-08). "Человеческая сборка dbSNP 150 удвоила количество записей RefSNP!". NCBI Insights. Получено 2017-05-16.
- ^ Jorde, LB; Вудинг, СП (2004). "Генетическая изменчивость, классификация и раса'". Природа Генетика. 36 (11с): S28–33. Дои:10,1038 / ng1435. PMID 15508000.
- ^ Тишкофф С.А.; Кидд, К.К. (2004). «Влияние биогеографии человеческих популяций на« расу »и медицину». Природа Генетика. 36 (11с): S21–7. Дои:10,1038 / ng1438. PMID 15507999.
- ^ Консорциум проекта «1000 геномов» (01.10.2015). «Глобальный справочник генетических вариаций человека». Природа. 526 (7571): 68–74. Bibcode:2015 Натур.526 ... 68 т. Дои:10.1038 / природа15393. ISSN 0028-0836. ЧВК 4750478. PMID 26432245.
- ^ Мондаль, Маюх; Бертранпедт, Жауме; Лео, Оскар (16 января 2019). «Приближенные байесовские вычисления с глубоким обучением поддерживают третью архаичную интрогрессию в Азии и Океании». Nature Communications. 10 (246): 246. Bibcode:2019НатКо..10..246M. Дои:10.1038 / s41467-018-08089-7. ЧВК 6335398. PMID 30651539.
- ^ Докрил, Питер (11 февраля 2019 г.). «Искусственный интеллект обнаружил в геноме человека неизвестного« призрачного »предка». ScienceAlert.com. Получено 11 февраля 2019.
- ^ Баррелл, Чирок (29 декабря 2019 г.). «Ученые внедрили ген человеческого интеллекта в обезьяну. Другие ученые обеспокоены». Обнаружить. Получено 30 декабря 2019.
- ^ Ши, Лэй; и другие. (27 марта 2019 г.). «Трансгенные макаки-резусы, несущие копии человеческого гена MCPH1, демонстрируют неотению развития мозга, подобную человеческой». Китайский национальный научный обзор. 6 (3): 480–493. Дои:10.1093 / nsr / nwz043. Получено 30 декабря 2019.
дальнейшее чтение
- Джоблинг, Марк А .; Холлокс, Эдвард; Херлз, Мэтью; Кивисилд, Тоомас; Тайлер-Смит, Крис (2013). Эволюционная генетика человека. Нью-Йорк: Наука Гарланд. ISBN 978-0-8153-4148-2. OCLC 829099073.
- Раннала Б., Ян З. (август 2003 г.). «Байесовская оценка времени расхождения видов и размеров предковой популяции с использованием последовательностей ДНК из нескольких локусов». Генетика. 164 (4): 1645–56. ЧВК 1462670. PMID 12930768.
внешняя ссылка
| Таксономия (Гоминины ) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Предки |
| ||||||||||||||||||||||||||
| Модели |
| ||||||||||||||||||||||||||
| Сроки | |||||||||||||||||||||||||||
| Другие |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| Подтемы | |
|---|---|
| Генетическая история по регионам | |
| Популяционная генетика по группе |
|
| |